Alguns aspectos da dinâmica da população dos hipopótamos caranx ( Peixes: Carangidae) em Ash Bocas, Caribe Colombiano
Alguns aspectos da dinâmica populacional dos hipopótamos de Crevelle Jack Caranx (Peixes: Carangidae) em Bocas de Ash, Caribe Colombiano
1ministração de agricultura e desenvolvimento rural, Bogotá, Colômbia.
2universidade da Magdalena, Programa de Engenharia das Pescas, Grupo de Biodiversidade e Ecologia Aplicada, Santa Marta, Magdalena, Colômbia.
3corporação Colômbia International (JRC), Barranquilla, Atlántico, Colômbia.
* Correspondência: [email protected]
recebido: setembro de 2009; Aceito: julho de 2010.
Resumo
Objetivo. O objetivo deste estudo foi avaliar aspectos biológicos e a dinâmica populacional dos ocos (caranx hipopótamos) no setor de Bocas de Ash, Caribe colombiano. Materiais e métodos. Entre junho de 2005 e junho de 2006, os dados de frequência de comprimentos comerciais foram coletados para determinar a relação de pulseira de pulso (LH-W), o tamanho médio da maturidade sexual (L50%), os parâmetros de crescimento (K, L ∞ e T0) taxas de mortalidade (m, f e z) e exploração (e). Havia 1151 indivíduos, dos quais 264 foram usados para análise biológica. Resultados. O rácio LH-W diferiu entre os sexos. Um L50% foi estimado em 63,6 cm de lh. Os parâmetros de crescimento apresentaram os seguintes valores: l∞ = 91 cm de lh, k = 0,38 ano-1 e = 0,32 ano. O valor do índice de desempenho teve um intervalo entre 3,33 e 3.73. A taxa de mortalidade total (Z) foi de 1,85 ano-1, mortalidade de pesca (f) de 0,72 ano-1 e natural (m) a 1,13 ano-1. O status de exploração (E) foi de 0,30 ano-1. Conclusões De acordo com a taxa de exploração, o jurado não apresenta superexploração em bocas de cinzas. No entanto, ao comparar o tamanho médio de captura com L50%, a espécie está sendo capturada com a principal engrenagem de pesca antes de 50% da população consegue amadurecer. Diretrizes para o desenho de um sistema de pesca que garanta a conservação e uso sustentável deste recurso.
Palavras-chave: Pesca, Reprodução, Caranx Hippos, Caribe, Colômbia. (Fonte: objetivos).
abstrato
Objetivo. Avaliar os aspectos biológicos e a dinâmica populacional do Crevelle Jack (Caranx Hippos) de Bocas de Ash, Caribe Colombiano. Materiais e métodos. Entre entre junho de 2005 e os dados de frequência de comprimento de junho de 2006 foram coletados para determinar a relação de peso do comprimento do garfo, tamanho médio de maturidade (L50%), parâmetros de crescimento da equação de von Bertalanffy (K, L∞ e T0), mortalidade (m, f e z) e Exploração (e) taxas. O total de 1151 indivíduos foram amostra, e 264 foram usados para análise biológica. Resultados O relacionamento de comprimento do garfo mostrou diferenças entre os sexos. L50% foi estabelecido em 63,6 cm. Os valores dos parâmetros de crescimento foram: l∞ = 91 cm fl, k = 0,38 ano-1 e = 0,32 ano. Os valores do índice de crescimento comparativo (φ ‘) variaram de 3,33 a 3.73. A taxa de mortalidade total (Z) foi estimada em 1,85 ano-1, mortalidade de pesca (f) como 0,72 anos-1 e mortalidade natural (m) como 1,13 ano-1. A taxa de exploração (E) foi determinada como 0,31 ano-1. Conclusões A taxa de exploração indicou que o Jack de Crevelle de Ash Bocas não foi superexplorado; Antes de 50% da população atinge sua maturidade sexual. Propomos ao enquadramento para o sistema de gestão da pesca, assegurando a conservação e uso adequado deste recurso.
Palavras-chave: Pesca, Reprodução, Hipopótamos de Caranx, Caribe, Colômbia. (Fonte: Objetivos).
Introdução
A família Carangidae é um dos grupos de peixes de maior importância económica nas pescarias tropicais e subtropicais do mundo (1, 2). De pouso da pesca artesanal no departamento Atlântico, os hipopótamos Caranx (Linnae 1766) é uma das espécies de maior demanda (3). Representa 47% em capturas desembarcadas anualmente e 70% da renda econômica da população de pesca, sendo capturadas principalmente com a transmissão e em alguns casos com ganchos e boliche (3). Apesar disso, há poucas informações detalhadas sobre os aspectos biológicos e populacionais que permitem elucidar o estado de exploração em que sua população é encontrada na foz do rio Magdalena.
As estimativas dos parâmetros de crescimento, mortalidade e taxas de operação são uma parte importante no estudo da biologia de peixe comercial, pois permitem determinar os níveis de exploração e fornecem ferramentas para a implementação de políticas de administração e controle para exploração adequada. (4-6). O objetivo deste trabalho foi determinar o rácio LH-W, o tamanho médio da maturidade sexual, os parâmetros de crescimento, as taxas de mortalidade e exploração de C. Hipopótamos em Ash Bocas para conhecer o estado de exploração neste setor e instrumento a científica Bases para propor diretrizes orientadas ao desenho de sistemas de pesca que levam a uma exploração sustentável desse recurso importante.
Métodos e métodos
Área de estudo. O estuário do rio Magdalena entendeu no Mar do Caribe, setor chamado Ash Bocas (11º 06 ‘- 11 ° 08’ ao norte; 74 ° 50 ‘- 74 ° 52’ Oeste; Figura 1). Este setor está localizado ao norte do Departamento Atlântico (3). O clima é caracterizado pela presença de dois períodos climáticos que afetam a atividade dos pescadores (7); O seco que começa no início de dezembro e termina no final de abril sem chuva e uma diminuição no fluxo do rio Magdalena com uma média de 4068 m3 s-1 (7, 8). O período chuvoso começa em maio e termina no início de dezembro com uma diminuição nos ventos e alta nebulosidade (7, 9) e é quando a maior água descarga com uma média de 10287 m3 s-1 (8) é apresentado.
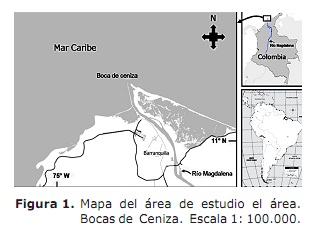
Bocas de Ash é um estuário de solução salina, onde sua operação responde à interação entre a onda maré e a contribuição fluvial dentro os contornos impostos pela geometria do mesmo estuário (9). O setor é influenciado por duas correntes: o Caribe do Caribe com um sentido oeste e a corrente do Panamá com isso (9). Nesta área, a atividade de pesca é tipo artesanal e tem cerca de 250 pescadores que operam as artes de pesca, ganchos, luminosos e artes de boliche. As capturas são representadas por espécies marinhas, bem como stuing espécies. Macrodon Ancyclodon, Caranx Crysos, C. Hipopótamos, Scomberomorus Braziliensis, Ariópis SP, SPP, SPP. e Arius Adereços (3).
Faixa de campo e laboratório. Entre junho de 2005 e junho de 2006, foram realizadas amostras por cinco dias de cada mês para registrar frequências de comprimentos de capturas comerciais pousadas no bairro de Las Flores, Barranquilla. As capturas foram realizadas com ganchos de 7-10 e com tigelas e transmeMágua entre 8,89 e 20,32 cm de tamanho de malha. Cada indivíduo era sexual e pesado (sem evisceração) com um saldo de precisão de 0,1 g. Como nem todos os peixes vieram com barbatanas completas, o comprimento do garfo foi usado. As medições foram feitas no limite inferior, usando um ictiômetro com precisão de 0,1 cm (3). Para a análise biológica, foram realizadas observações macroscópicas das gônadas para determinar o sexo e o estado de maturação gonadal, tomando como referência a classificação qualitativa de Vazzoleler (10), que consiste em quatro estados: indiferenciados; imaturo (i); Na maturação (II); Maduro (iii) e gerado (iv).
taxa de comprimento (LH-W). A relação de comprimento-peso foi determinada por sexo através da equação potencial w = ALHB, onde W é o peso total, A e B são constantes da regressão potencial e LH é o comprimento do garfo. Para determinar as diferenças no rácio LH-W entre os sexos, foi realizada uma análise de covariância (ANCOVA), onde a variável dependente era W, a variável independente foi lh e o fator sexual. Antes da análise, os pressupostos da normalidade foram avaliados (Kolmogorov-Smirnov), homogeneidade de variações (teste Bartlett), relação linear entre a variável W e LH (análise de correlação) e homogeneidade dos brincos da relação entre W e LH .
Meio estoque de maturidade sexual (L50%), composição de captura por tamanhos e tamanho médio de captura (TMC). Avaliar o impacto da pesca na biologia de C. Hipopótamos, L50% foi calculado. Esses espécimes cujas gônadas foram observadas nos Estados III e IV foram consideradas. O cálculo foi baseado no modelo de logística que relaciona a proporção de indivíduos maduros com comprimento (l; 5):
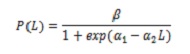
, β = 1; α1 e α2 são os parâmetros da regressão.
Estimar L50%, a proporção -α1 / α2 foi utilizada. Para ajustar o modelo, o método de regressão não linear foi usado por quadrados mínimos, usando o Algoritmo de Newton iterativo.
A composição da captura de tamanho foi usada para calcular o TMC e os parâmetros de crescimento.Os dados foram agrupados mensalmente em colegas a cada dois centímetros. A relação entre a frequência relativa e o número total de indivíduos foram utilizados para calcular o TMC usando o procedimento proposto por Sparre e Venema (5); Posteriormente, um TMC foi calculado para cada arte de pesca observada. O TMC foi comparado com L50% para avaliar o impacto da pesca na biologia da espécie.
Parâmetros de crescimento. Os dados de comprimento foram agrupados mensalmente em colegas de 1 cm. Os parâmetros de crescimento foram estimados por três rotinas contidas na versão 1.1 (4) do FISAT Computacional: a análise de frequência eletrônica (Elefan I), a de comprimentos de folha de pastor (SLCA; 11) e o método de projeção matrica (Projmat; 12) . A rotina Elefan i trabalha com a equação de crescimento sazonalizada de von Bertalanffy (Elefan I), sendo o tamanho expresso de acordo com a idade (5):
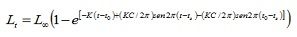
onde, lt é o comprimento para a idade t; L∞ o comprimento assintótico; K é a constante de crescimento, para o parâmetro de condição inicial, que determina o ponto hipotético no tempo em que o peixe tem um tamanho igual a zero; C é o parâmetro de amplitude da oscilação do crescimento; TS faz parte do ano em que a taxa de crescimento é mínima.
A rotina do SLCA estima o crescimento independente do número e da posição das modas na amostra, com base na configuração de função cosseno:
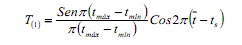
onde, tmax e tmin são as idades correspondentes aos limites superior e inferior de um determinado intervalo de classe; É a idade média e TS é a fração do ano em que a amostra foi retirada (6).
O projmat projeta uma amostra de distribuição de frequência do tempo T + 1, usando o K e o L∞ valores. As frequências projetadas foram comparadas com as observadas pela soma das diferenças de quadrados, repetindo o processo até obter as melhores estimativas (6).
Levando em conta que não é possível obter uma estimativa para a partir das frequências de comprimentos (13,14), este parâmetro foi estimado a partir da equação empírica de Pauly (15):
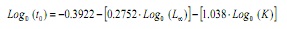
Onde é a idade hipotética à qual o animal começa a crescer, l∞ o comprimento assintótico e k é a constante do crescimento
Comparar estimativas de crescimento entre os métodos, foi utilizada a taxa de desempenho de crescimento φ ‘(16). Além disso, esses valores foram comparados com as estimativas de outras populações de C. hipopótamos documentados na literatura. Este índice foi estimado a partir da expressão:
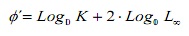
O que, k e l∞ são os parâmetros de crescimento.
Dado que o valor estimado de L∞ neste estudo foi baseado em lh e aqueles documentados na literatura, em total comprimento (LT), para fins de fazer comparações entre os valores, uma simples regressão linear foi feita entre o LT (dependente) e lh (independente) para tornar a conversão de lh para lt. Para isso, o seguinte modelo linear foi obtido com dados deste estudo:
lt = – 0,4387 + (1.2335 lh)
das freqüências de lh de capturas comerciais, a mortalidade total (Z), natural (m) e pesca (f) da população de C. hipopótamos voltou a estimar. Z foi estimado a partir da curva de captura linearizada convertida para tamanhos (17):
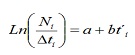
Onde, nem o número de indivíduos capturados pertencente à classe de comprimento i; T’i é a adega relativa da classe I; Δti é o tempo médio exigido pelo indivíduo por seu crescimento através da classe I.
m foi estimado usando a equação empírica de paully (17):
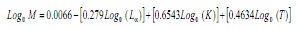
Onde, l∞ e k são os parâmetros da equação von Bertalanffy e T é a temperatura média anual da água costeira superficial na área de estudo, que foi considerada a 28,2 ° C. / p>
A taxa de mortalidade de pesca e a taxa de operação foram estimadas a partir de:
Para executar as estimativas dos parâmetros, foi assumido um crescimento sazonalizado de C. hipopótamos, dado que ao longo do ano, episódios ambientais na área de estudo que podem estar influenciando o crescimento dos peixes. Tal é o caso das condições sazonais do caudal do rio Magdalena, que pode influenciar as condições físicas, químicas e biológicas em sua boca (8). Para isso é adicionado, a influência dos ventos de alium no setor da boca de Ash, uma vez que quando estão em andamento e de acordo com sua intensidade, eles também mudam as condições oceanográficas do setor (18). Da mesma forma, o conhecimento biológico da espécie indicam que C.Os hipopótamos são uma espécie migratória (1), da qual a sazonalidade também pode ser inferida em crescimento. Com base no acima, os valores encontrados por Elefan I foram selecionados para estimar as taxas de mortalidade de C. Hipopótamos neste estudo.
Com o objetivo de quantificar o efeito das amostras mensais nos parâmetros de crescimento e Obtenha os valores de precisão do mesmo, a técnica de remampamento JackKnife (6) foi aplicada. Isso consiste em estimar os tamanhos mensais de frequência mensal e seqüencialmente são omitidos para obter combinações de cada um dos parâmetros. Para especificar esta estimativa, foi utilizado o erro percentual (EP) e o coeficiente de variação (CV), utilizando as seguintes fórmulas:
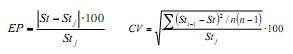
Onde, St é o parâmetro estimado de crescimento usando o banco de dados anual completo e o STJ é o estimador médio derivado das estimativas sucessivas do JackKnife; O STI-1 é o parâmetro estimado ao omitir o mês I, e N é o tamanho da amostra anual (n = 12).
Resultados
Durante o período de estudo 1151 indivíduos foram coletados, dos quais 868 foram capturados com Transmal, 120 com um gancho e 167 com boliche; Apresentando uma gama de tamanho entre 10,5 e 96,5 cm de comprimento. Para a análise dos aspectos biológicos estudados, 264 indivíduos foram contados, sendo 180 homens e 84 fêmeas, com uma gama de tamanho entre 25 e 96,5 cm de LH e um peso total entre 281,5 e 10500 g.
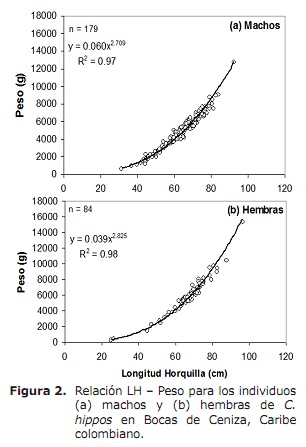
taxa de comprimento (LH-W). A análise revelou que entre o sexo o rácio LH-W diferiu significativamente, observando que as fêmeas pesam mais do que os machos no mesmo comprimento (Ancova: F1, 260 = 21,5, p < 0,05 ). Portanto, a relação LH-W para homens foi W = 0,060LH2.709 (p < 0.05) e para as fêmeas w = 0,039LH2.825 (p < 0,05) (Figura 2). Os valores de B resultantes de regressões para ambos os sexos foram significativamente inferiores a 3, indicando um crescimento menor (teste t, p > 0,05; 15).
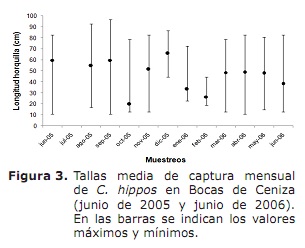
Tamanho do tamanho, Tamanho médio do vencimento sexual (L50%) e tamanho médio de captura (TMC). Em geral, os tamanhos de captura mínima foram registrados em outubro de 2005 e março, abril e junho de 2006; Enquanto as máximas eram frequentes durante todo o período do estudo, exceto para outubro, janeiro e fevereiro (Figura 3).
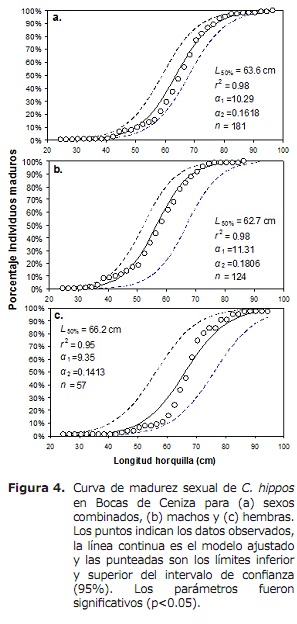
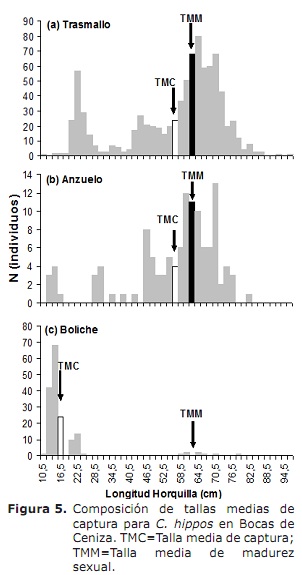
l50% para sexos combinados foi de 63,6 cm LH (95% CI: 59,0-68,4 cm). Para o caso de machos e fêmeas foram 62,6 cm (95% CI: 58,3-70 cm) e 66,2 cm (IC 95%: 56,1-76,2 cm), respectivamente (Figura 4). O TMC global foi estimado em 50,3 cm de lh. Para o caso de transmmas, ganchos e boliche foram estimados em 56,0, 56,2 e 17,8 cm, respectivamente (Figura 5A, 5B e 5C), abaixo de L50%.
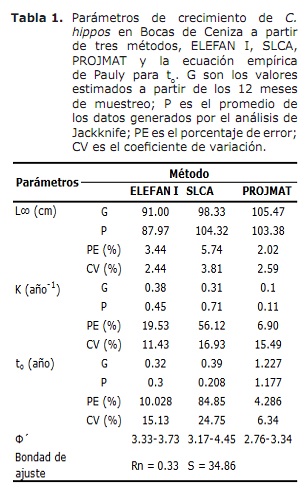
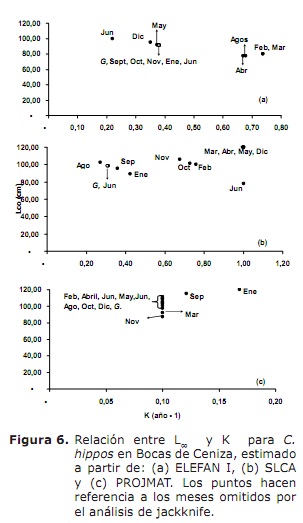
crescimento. Os resultados de L∞ e K fornecidos pelos métodos Elefan I, SLCA e Projmat são apresentados na Tabela 1. Observa-se que esses três métodos forneciam valores diferentes para cada um dos parâmetros de crescimento. A análise de JackKnife mostrou que há um efeito dos dados sobre os diferentes parâmetros de crescimento, especialmente para o SLCA e o Projmat. Para o caso de Elefan I, L∞ variei de 77,4 a 100 cm, enquanto os valores de K estavam entre 0,20 e 0,75 ano-1 (Figura 6). Com o SLCA, os maiores valores de L∞ e K foram obtidos com valores entre 78,25 e 120 cm e entre 0,3 e 1,0 ano-1, respectivamente. De acordo com os valores lançados pelo Projmat, menos dispersão foi observada para K flutuando entre 0,10 e 0,17 ano-1. O oposto ocorreu para L∞, onde os valores variaram entre 87 e 120 cm, sendo o último maior do que o observado nos estimadores de crescimento anteriores. Os valores de K estimados mostraram diferenças entre os métodos, com os mais altos calculados por Elefan I e SLCA (Tabela 1). O valor obtido para a idade hipotética a que o animal (a) começa a crescer pela equação empírica de Paully (20) foi de 0,32 anos, com um coeficiente de variação de 13,92% e uma porcentagem de erro (PE) de 10.02. A análise do índice de desempenho ǿ mostrou valores semelhantes entre elefan I (3.33-3.73) e SLCA (3.17-4.45), que não ocorreu para projmat que mostrou valores inferiores aos anteriores (2.76-3.34). Mesmo assim, a menor variação foi apresentada para Elefan I, seguido por Projmat e SLCA, sendo este último a mais alta classificação apresentada. Tendo em conta o coeficiente de variação (CV) de cada um dos parâmetros, descobriu-se que o Elefan I estima da análise de JackKnife tendia a fornecer valores mais baixos para L∞, ao contrário do SLCA e ao Projmat que atingiram os valores mais altos (Tabela 1) .O resultado do PE obtido para k com projmat foi menos do que o encontrado para Elefan I e SLCA. Por conseguinte, o método que lançou as estimativas mais credíveis dos parâmetros de crescimento foi o Projmat, seguido por Elefan I e Finalmente SLCA.
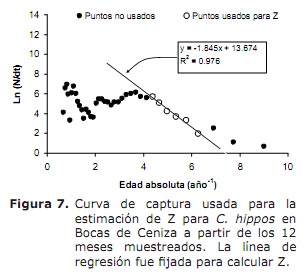
Mortalidade e status de exploração. Com base nos seguintes parâmetros de crescimento: K = 0,38 ano-1; L∞ = 91 cm lh; para = 0,32 anos; Oscilação sazonal no padrão de crescimento (c) de 0,5 e taxa de crescimento mínima (WP) de 0,3, uma taxa de mortalidade total (z) de 1,76 ano-1 (Figura 7) foi estimada, oscilando entre 1,38 e 2,22 anos -1 ( da análise de Jackknife).
Mortalidade natural (m) foi de 1,12 anos-1, variando entre 1,10 e 1,14 ano-1. A taxa de mortalidade de pesca (F) foi estimada em 0,63 ano-1, apresentando uma gama de variação entre 0,2 e 1,06 anos-1. Os valores da relação F e Z para estimar a taxa de operação (E), apresentaram um valor máximo de 0,5 ano-1, uma média de 0,30 ano-1.
Discussão
taxa de comprimento (LH-W). O conceito de crescimento envolve mudanças relativas no tamanho e no peso dos indivíduos (19), e sua relação tem vários usos na pesquisa de pesca. Por exemplo, é útil nos modelos de análise de estoque de pesca para estimar a biomassa de uma população; conhecer a condição de um peixe; comparar a história da vida de certas espécies entre as regiões; e determinar o tipo de crescimento que os peixes geralmente presentes (20). C. Hipopótamos, apresenta um peso livre de peso do tipo alométrico (< 3; 15), indicando que apresenta taxas de crescimento diferenciais para diferentes partes do corpo, onde o comprimento aumenta a um taxa mais alta que o peso.
Os resultados indicam que esta condição foi mais evidente para os homens do que para as fêmeas, dado que este último, apesar de apresentar um menor crescimento alométrico, aproximado para ter crescimento isométrico. Isso foi concordante com o resultado Ancova, que indicava que o mesmo LH há diferenças no peso sexual. As fêmeas desta espécie apresenta maior peso e altura do corpo do que os machos. Por outro lado, é importante destacar que, porque as amostras foram coletadas por um ano, a relação LH-W neste estudo não representa uma época particular do ano. Nesse sentido, os valores B podem ser considerados como um valor médio anual para as espécies (20). Os resultados desta pesquisa foram semelhantes aos documentados para as espécies em diferentes áreas geográficas. Por exemplo, os valores B baseados em B foram documentados de 2.73 no sul da Flórida (21), de 2,85 na Índia (1) e 2,91 no Golfo de Salamanca, Colômbia (22).
Crescimento, estado de mortalidade e exploração. Com base nos parâmetros de crescimento obtidos por Elefan I, o comprimento máximo estimado para C. hipopótamos em Ash BOCAS foi de 91 cm LH correspondente a 111,81 cm de LT. Se este último for convertido para a idade correspondente a 14 anos, obtido com uma taxa de crescimento K de 0,38 ano-1, sugerindo que C. hipopótamos é uma espécie de Longeva (17). Por outro lado, levando em conta a curva de crescimento, esta espécie cresceu rapidamente até os 5 anos de idade. Esses dados coincidem para outros arquivamentos, como o de Trachinotus falcatus, que atinge um tamanho máximo de 90 cm e também tem um crescimento máximo de 5 anos, da qual inicia a fase estacionária de seu crescimento (2).
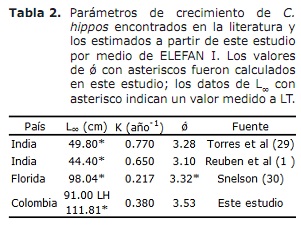
Por outro lado, os valores de L∞ estimados para os hipopótamos em C. neste estudo foram superiores aos documentados na literatura; Enquanto os do Índice de Desempenho ǿ revisados na literatura mostraram valores muito próximos da estimativa neste estudo (Tabela 2). Tomando este índice como um critério de comparação entre os valores dos parâmetros de crescimento obtidos e tendo em conta que C. hipopótamos é uma espécie migratória (23), pode-se assumir que o crescimento desta espécie é concordante naquela descrita em Diferentes áreas de sua distribuição global (Tabela 2).
O padrão de crescimento anual de C. Hipopótamos mostrou oscilações (c = 0,5) com uma taxa mínima de crescimento no mês de setembro (WP = 0,3). Este padrão pode ser influenciado por um dos aspectos mais característicos do crescimento dos peixes: a variabilidade observada entre indivíduos de uma população e entre as populações de uma espécie, que está diretamente relacionada ao meio ambiente (24). Por exemplo, a variabilidade nas condições físico-químicas da água (24) e a qualidade e a quantidade de alimentos (19) condicionam a migração de uma espécie, que engloba deslocamentos de estação entre as zonas de criação, alimentação e desova (15, 25) .
com base no acima e tendo em conta as frequências de comprimentos observadas durante o período de estudo, que também indicam a possível presença de sazonalidade no crescimento (13), pode-se inferir que o crescimento de C. hipopótamos é sazonal. Em relação à taxa de crescimento mínimo, este evento pode ser dado pelo estágio reprodutivo das espécies, já que foi observado um aumento no número de indivíduos maduros (em etapas III).
Por outro lado, a relação entre a taxa de mortalidade de pesca (F) e a mortalidade total (z) de 0,3, indica que a população na área de bocas de cinzas não é correr o risco de ser superexplorado , sob a premissa de que uma população atinge seu desempenho ideal quando a fração de morte causada por F está acima de 0,5 (6,13). No entanto, é importante destacar que, quando L50% foi comparado ao TMC, C. hipopótamos apresentou maior pressão sobre a população adulta, freqüentemente capturada com transmissão e gancho. A situação piora com o uso de Boliche, que não só afeta adultos, mas também os jovens, o que não permite a renovação natural desse recurso de pesca.
Proposta para o desenho de um sistema de manuseio de pesca . A partir da base de que a sobrepesca pode ocorrer de três maneiras diferentes: (i) em relação ao crescimento; (ii) em relação ao recrutamento; e (iii) a sobrecarga que afeta o ecossistema (15). Nesse sentido, a pesca de C. Hipopótamos no setor da boca de Ash pode ser classificada dentro do primeiro item, que ocorre quando os indivíduos são capturados antes de alcançar o tamanho necessário para garantir a renovação biológica da população. O acima se tornou um problema comum para a maioria das espécies comerciais, e foi documentado para muitas espécies costeiras no Caribe colombiano.
Todos os itens acima, é necessário que os esforços estejam investindo para projetar gerenciamento de estratégias para C. Os hipopótamos na pesca de Bocks Asho, de tal forma que sejam consistentes com a abordagem de precaução para a pesca responsável por este recurso (27). Caddy (27) sugere que as estratégias devem ser aplicadas dentro de uma estrutura de redundância de gestão em que tal estrutura poderia consistir em uma mistura de medidas envolvendo artes, critérios de mergulho temporário e aumento de tamanhos mínimos de captura. Nesse sentido, levando em conta o L50% como ponto de referência limite (28), é recomendável que o tamanho mínimo de captura deste recurso seja fixado a 64 cm de comprimento. Da mesma forma, em todas as artes de pesca utilizadas, propõe-se aumentar os tamanhos de malha acima de 8,89 cm e usar calibres de gancho destinados a capturar peixes maiores. Nas condições atuais, essas engrenagens de pesca estão afetando diretamente a fração de jovens que entram na primeira fase de maturidade sexual. Com as medidas acima, complementadas com outras variáveis, como captura, o esforço de pesca e captura por unidade de esforço, bem como fatores sociais e econômicos, espera-se que C. hipopótamos seja capturado em uma maior proporção de indivíduos acima de L50% para garantir a sustentabilidade desse recurso ao longo do tempo. Nesse sentido, é importante levar em conta o status multipecífico e multiflot da pescaria e, portanto, recomenda-se um processo de concordância entre os pescadores, os comerciantes e as entidades relevantes para atingir acordos destinados a uma exploração responsável do recurso.
Em conclusão, o jorel não apresenta superexploração em bocas de cinzas. No entanto, ao comparar o tamanho médio de captura com L50%, a espécie está sendo capturada com a principal engrenagem de pesca antes de 50% da população consegue amadurecer. Diretrizes para o desenho de um sistema de pesca que garanta a conservação e uso sustentável deste recurso.
Reconhecimentos
para a comunidade dos pescadores, membros do Asopscar e COOPEZ. Para os irmãos Villa e La Pescadería El Nuevo Tambó pela sua colaboração na coordenação de atividades e fornecimento de informações de campo. Para os professores G. Lozano, l.c. Gutiérrez, O. Galvis, J. White e E. Viloria.
referências
1.reuben, S, HM Kasim, S Sivakami, PN Radhakrihhnan, Kurup, Kurup, M sivila, nobre, kvs nair, raje sg. Pesca, biologia e avaliação de estoque de recursos carangur dos mares indianos. Indian J Fish 1992; 39 (3,4): 195-234.
2.crabtree, R, PB Hood, Snodgrass d. idade, grouth e reprodução de permissão (trachinotus falcatus) nas águas da Flórida. Fish Bull 2002; 100: 26-34.
3.River R, Solano, G.Diagnóstico da atividade de pesca artesanal na área costeira do Departamento Atlântico, Caribe colombiano. Barranquilla, Colômbia: Incoder; 2005.
4.Gayalino, FC, P Sparre, Paully D. O guia do usuário do FISAT. FAO – Ferramentas de asessment de estoque de iclarm. Roma: FAO; 1993.
5.SPARRE, P., VENEMA S. Introdução à avaliação dos recursos pescadores tropicais. Roma: FAO; 1995. Documento de pesca técnica: 306/1.
6.Rueda, M, Santos-Martínez A. Dinâmica populacional do Listrado Mujarra Eu Lmerieri do Big Ciénaga de Santa Marta, Colômbia. Fish Res 1999; 42: 155-166.
7.blanco, JA, EA Viloria, Narváez BJC. ENSO e mudanças de salinidade interunnais no sistema de lagoa costeira do Big Ciénaga de Santa Marta, Caribe colombiano. Estuar Costa Shelf Sci 2006; 66: 157-167.
8. Restrepo, JD, Kjerfve B. Magdalena River: variabilidade interunnal (1975-1995) e estimativas revisadas de descarga de água e carga de sedimentos. J hidrol 2000; 235 (1-2): 37-149.
9.acevedo A, Gutiérrez ml. Alguns aspectos reprodutivos de Rhizotrionodon Toyo (1831) poroso (elasmobranchii: Carccharinidae). No setor da boca de Ash, Caribe colombiano. . Barranquilla, Colômbia: Universidade Atlântica, Faculdade de Ciências Básicas; 1999.
10.Vazzoleler, EAM. Método Manual para Peixes Popleare Biological Studists: Reprodução e Crescimento. Brasília: CNPq; 1982.
11.Shoperd, JG. Um método fracamente paramétrico para a análise de dados de composição de comprimento. Em: Paully, D, editores de Morgan GP. Métodos baseados em comprimento na pesquisa das pescas. Manila, processo de conferência ICLarm 13. 1987.
12.Lielie, pH. O uso de matrizes em certas matemáticas populacionais. Biometrika 1945; 35: 213-245.
13.gulland, ja, rosemberg AA. Exame dos métodos baseados no tamanho para avaliar as populações de peixe. Roma: FAO; 1992.
14.Tordecilla-Petro G, S Sánchez-Band, Olaya-Nieto Ch. Crescimento Moncholo e Mortalidade (Hoplias Malabaricus) no Lorica Lorica, Colômbia. Rev MVZ Córdoba 2005; 10 (2): 623-632.
15.pauly, D. Alguns métodos simples para a avaliação dos recursos pescadores tropicais. Roma: FAO; 1983.
16.pauly, D, Munro JL. Onze mais sobre o comboio de crescimento em peixes e invertebrados. Fishb 1984; 1:21.
17.pauly, D. Dinâmica da população de peixes em águas tropicais; Um manual para uso com calculadoras programáveis. Prisioneiro ICLarm Rev 1984; 8: 235.
18.blanco, ja. Variações ambientais sazonais nas águas costeiras e sua importância para a pesca na região do Caribe de Santa Marta, da Colombiana. . Santa Marta-Bogotá. Universidade Nacional da Colômbia, Faculdade de Ciências Básicas; 1988.
19.abaunza L, Karlou-Riga C, Murta a, Eltink W, García Santamaría Mt, Zimmermman C, Martelo C, et al. Crescimento e reprodução da cavala do cavalo, trachurus trachurus (Carangidae). Rev Fish Biol Fish 2003; 13: 27-61.
20.Froese, R. Lei de Cubo, Fator de Condição e Relacionamento de Peso: História, Meta-Análise e Recomendações. J appl ichthyol 2006; 22: 241-253.
21.bohsack, ja, harper de. Relacionamentos de peso de comprimento de peixes de recife marinhos selecionados do sudeste dos Estados Unidos e do Caribe. Memorando técnico noaa; 1988. NMFS-SEFC-215.
22.Duarte, Lo, CB García, I Moreno, G Melo, P Navajas, N Sandoval, et al. Atlas demográficas do Golfo de Salamanca, Caribe Colombiano: Dinâmica População, Distribuição, Comida e Reprodução. . Versão 1. Santa Marta: invema-unal-colciencias; 1999.
23.HoneyBrink, R. Uma revisão da biologia da família Carangidae Whith Ênfassis nas espécies encontradas em águas havaianas. Honolulu: divisão de recursos acuáticos; 2000.
24.Macer, CT. Alguns aspectos da biologia da caverela do cavalo (trachurus trachurus (L)) nas águas ao redor da Grã-Bretanha. J peixe biol 1977; 10: 51-62.
25.Harden, fr. Migração de peixes. Londres: Arnold; 1968.
26.Narváez B, JC, F Herrera, White J. Efeitos do equipamento de pesca no tamanho do peixe em uma pesca artesanal caribenha colombiana. Bol investir Mar Mar 2008; 37 (2): 163-187.
27.Caddy, JF. Gestão das pescas no século XXI: serão aplicáveis novos paradigmas. Rev Fish Biol Fish 1999; 9: 1-43.
28.Caddy, JF. Limite de pontos de referência, semáforos e abordagens holísticas para a gestão das pescas com o mínimo de avaliação de estoque. Fish Res 2002; 56 (2): 133-137.
29.Torres, FJR, MBTM Norizam, LR Garcés, Silvestre Gt. Compillation de parâmetros de população de espécies de peixes comumente pegos em arraswls no sul e sudeste da Ásia. Manila: World Fish Center; 2001.
30.Snelson, FF. Estudos biológicos de Crevelle Jack (Hipopótamos Caranx) na Flórida. São Petesburg: Instituto de Reseach Marine Florida; 1992.