L’expression de l’IL-10 intervient dans la régulation de la réponse inflammatoire par oméga 3 acides gras
s. Sierra, F. Lara-Villoslada, M. Olivares, J. Jiménez, J. Boza et J. Xaus
Département d’immunologie et d’études précliniques. PULEVA BIOTECH. Grenade.
Résumé
Cible ou arrière-plan: Acides Polyunosaturé Les graisseuses sont importantes pour l’organisme humain en raison de sa participation à de nombreuses fonctions biologiques. Les régimes occidentaux se caractérisent par d’être excessivement riches en acides gras W-6 et pauvres en acides gras W-3. Les acides gras de la série W-3 sont nécessaires à la croissance et au développement normaux de l’individu ainsi que pour la réglementation de la réponse immunologique. L’objectif de cette étude est d’analyser l’effet d’un régime enrichi en acides gras W-3 contre un processus inflammatoire ainsi que de l’étude des mécanismes impliqués dans ledit effet. Interventions: Pour ce faire, les souris Balb / C ont été nourries pendant un mois avec un régime alimentaire dont la source de lipides était de 100% d’huile de tournesol (contrôle) ou avec le même régime dans lequel 12% de la graisse était de l’huile de poisson et de la Huile de tournesol à repos (W-3). Douze heures avant que son sacrifice ne soit induite dans l’une des oreilles de chaque animal une dermatite de contact qu’il a étudié avec une inflammation et un œdème. En tant qu’agent inflammatoire, 2,4 dinitrofluorobenzène ont été utilisés. Après le sacrifice, divers échantillons ont été pris et analysés. Résultats: une inflammation, mesurée comme une teneur en poids et en eau des oreilles, a diminué de manière significative chez les souris propulsées par W-3. La mesure de l’infiltration de leucocytes et des paramètres d’oxydation ont également révélé l’amélioration du processus inflammatoire desdites RA-TONES. Pour expliquer ces faits, l’expression de divers cytokines a été analysée, une augmentation de l’IL-10 et une diminution des cytokines TH1 et TH2 ont été analysées. Conclusions: Les acides gras G-3 ont un effet immunomodulateur lorsqu’ils agissent comme anti-inflammatoires et anti-allergies, tout en augmentant certaines défenses de l’organisme. La cytokine réglementaire IL-10 pourrait être responsable de l’effet anti-inflammatoire exercé par les acides gras W-3.
L’expression de l’IL-10 est indolée dans la régulation De la réponse immunitaire des acides gras oméga-3
abstrait
Introduction: Les acides gras polyinsaturrarés jouent un rôle clé dans un grand nombre de fonctions biologiques. Les régimes occidentaux sont très riches dans les acides gras W-6. Cependant, la teneur en acides gras W-3 ne convient pas dans ces régimes, le déshonquent de leur importance dans le développement normal du corps humain et de la réglementation de la réponse immunitaire. Le but de ce travail est d’examiner l’effet de l’alimentation enrichie des acides gras W-3 dans la régulation de la réponse inflammatoire. Matériel et méthodes: Les souris BALB / C ont été nourries à l’aide de l’alimentation riche en acides gras W-6 (huile de tournesol 100%) ou au régime fortifié de l’acides gras W-3 (12% d’huile de poisson plus de 88% d’huile de tournesol) pendant 28 jours. Douze heures avant le sacrifice, les souris ont été traitées avec le 2,4-ninitro-1-fluorobénène sur l’oreille gauche pour induire la réaction inflammatoire. Ensuite, les souris ont été sacrifiées et les différents échantillons collectés ont été analysés. Résultats: L’oreille inflammation des souris nourries Le régime W-3 était nettement inférieur. L’infiltration des leucocytes et le stress oxydatif étaient également plus bas dans ces souris. Pour expliquer ces résultats, une expression de cytokine et une concentration en eicosanoïde plasmatique ont été mesurées. Une augmentation des niveaux IL-10 et une réglementation en baisse des réponses TH1 et TH2 ont été observées chez les souris nourries au régime W-3. Conclusion: non seulement les acides gras n-3 exerce un rôle antiinflamatoire et un rôle antialergique, mais elles améliorent également certaines des défenses de l’organisme. Nos données suggèrent que les acides gras W-3 réduisent régulent la réponse inflammatoire en améliorant l’expression IL10.
(Nut-HOP 2004, 19: 376-382)
Mots clés: Acides gras polyinsaturés. Anti-inflammatoire Organisme.
Correspondance: J. Xaus. Département d’immunologie et d’études précliniques. Puléva Biotech. Road de Purlu, 66. 18004 Grenade. E-mail: [email protected]
reçu: 21-I-2004. Accepté: 13-II-2004.
Introduction
Le système immunitaire est impliqué dans la défense de l’organisme contre des agents étrangers, des cellules tumorales et des agressions extérieures. Un grand nombre d’organes, de cellules, de tissus et de molécules interagissent pour mener à bien cette mission.
Le développement, la maintenance et le fonctionnement optimal du système immunitaire dépendent du régime entre autres facteurs. Parmi les nutriments qui affectent la plupart du système immunitaire met en évidence la graisse dont la carence ou la contribution déséquilibrée ainsi que la qualité de celle-ci compromet la stabilité du système immunitaire1.
Le rapport entre les acides gras et l’inflammation est principalement due à la composant phospholipide des cellules du système immunitaire. Les acides gras incorporés dans la membrane d’entre eux peuvent affecter les fonctions de celui-ci et donc du fonctionnement des cellules. Les phospholipides membranaires sont responsables de la fluidité de celui-ci, ce qui affectera les différents composants ancrés. Différents signaux intracellulaires quittent ces phospholipides et leur composition affecte donc différentes voies de transduction de signal. De plus, les acides gras synthétisés à partir de phospholipides sont des substrats de la synthèse d’un groupe de médiateurs, d’eicosanoïdes, d’une importance biologique marquée2.
Actuellement, l’intérêt de l’étude des acides gras est en augmentation et l’influence d’eux est important sur la réponse immunologique. Cet intérêt s’est concentré sur l’étude des acides gras polyinsaturés à longue chaîne (AGPI-CL), venant de familles W-6 et W-3, dont les précurseurs, les acides linoléiques et l’α-linolénique sont essentiels, à ingérer avec la le régime alimentaire, aussi bien qu’il est également recommandé d’inclure certains de ses dérivés, tels que l’acide eicosapentaénoïque (EPA), le docosahexaénoïque (DHA) et Arachidonic (AA), depuis les voies métaboliques, par allongement de ses précurseurs, les niveaux appropriés sont pas atteint3.
chez les personnes qui consomment des régimes occidentaux, les cellules inflammatoires contiennent des proportions élevées d’AGPI-CL de la série W-6, en particulier des proportions et de faibles proportions, de l’EPA de la W-34 séries. Le rôle principal de l’AA est d’agir en tant que substrat pour la synthèse de médiateurs bioactifs du type Eicosanoid. Ceux-ci sont impliqués dans la durée et l’intensité modulaires de la réponse inflammatoire. Les dérivés AA, tels que TXA2 et LTB4, sont des agents puissants avec un effet inflammatoire marqué. Le TXA2 dispose d’un vasoconstricteur et d’un effet proagrénique tandis que le LTB4 est un puissant agent chimiotactique5. De plus, le PGE2 induit la fièvre, augmente la perméabilité vasculaire et la vasodilatation, tandis que la production de cellules IgE b.
L’augmentation de la consommation d’acides gras W-3 entraîne une diminution de l’AA dans les membranes de Les cellules inflammatoires décroissant ainsi leur capacité à synthétiser de la COX et des eicosanoïdes dérivés de l’AA. En plus d’inhiber le métabolisme AA, l’APE inhibe l’oxygénation AA par Cox et supprime l’induction de certaines cytokines agissant sur l’expression génique de COX-2 et 5-LOX. Il est également capable d’agir en tant que substrat de Cox et 5-Lox, donnant lieu à des eucosanoïdes considérés comme biologiquement moins actifs5.
Tout cela conduit à l’idée que l’huile de poisson a des propriétés anti-inflammatoires. La consommation chez les individus en bonne santé d’une proportion adéquate des acides gras G-6 et W-3 entraînerait une meilleure réglementation de la réponse immunologique et que l’augmentation de la proportion d’acides gras W-3 serait bénéfique pour les sujets qui souffrez de maladies auto-immunes ou inflammatoires dues à l’effet anti-inflammatoire d’eux1.
Non seulement ces mécanismes sont impliqués dans l’effet anti-inflammatoire, mais leur implication de la voie de signalisation est en cours que NFKB, MAPK ou PPAR et lxr6. L’action à travers ces deux récepteurs ouvre un nouveau champ de recherche et une plus grande gamme d’applications des acides gras G-3 en raison de l’emplacement et de la fonction d’entre eux.
La présente étude consiste à démontrer l’anti- Effet inflammatoire des acides gras G-3 et des mécanismes impliqués, mesurant les paramètres immunologiques permettant de vérifier son effet au niveau systémique et d’analyser le cours d’une inflammation induite pour déterminer son niveau local.
matériau et méthodes
réactifs et anniversaires
Concanvalin A (avec a) et lipopolysaccharide (LPS) ont été fournis par Sigma Chemical Co (St Louis, MO). Les anticorps fournis par Bethyl (Montgomery TX) ont été utilisés pour mesurer les immunoglobulines et dans le cas de la cytokines par biosource (Camarillo, CA). Peroxydase radis, bromure d’hexadécyltriméthylammonium, chlorhydrate d’o-dianysidine et peroxyde d’hydrogène pour la mesure de l’activité de la milopétoxidase ainsi que l’acide trichloroacétique, la β-nadpa, le 5,5-ditio-bis (nitrobenzoïque à 2 acides) et le glutathione ont également été acquises à Sigma . La glutathione réductase a été fournie par Roche.L’eau désionisée purifiée a été utilisée avec un système Milli-Q (Bedford, MA).
Animaux et régimes
Les souris mâles Balb / C au sevrage de trois semaines ont été fournies par l’université. de Grenade. Ils ont été maintenus tout au long de l’expérience à une température contrôlée de 22 ° C et sous un cycle de lumière de 12 heures. L’expérience globale a été effectuée 3 fois avec des résultats similaires.
Toutes les études d’animaux effectuées ont été réalisées selon les normes d’éthique des animaux définies aux critères Helsinki.
L’expérience se compose de 2 groupes de souris (n = 10) alimentés pendant 28 jours avec des régimes égaux de base et dont la différence réside dans le type de graisse incluse. Les deux contiennent 5% de graisse de régime total. Cette graisse de 5% est constituée d’une huile de tournesol dans le cas du groupe utilisé comme contrôle. Dans le cas du régime alimentaire W-3, 12% de la teneur en graisse totale est une huile de poisson raffinée, tandis que le reste de la graisse est la même huile de tournesol du groupe précédent. Les deux régimes ont été élaborés à partir de la mise en place spécifique à la suite des recommandations de l’Académie vétérinaire américaine pour la nutrition des animaux. De cette manière, des régimes semi-synthétiques ont été préparés comprenant 5% de graisse, 21,3% de protéines, de 67,5% de glucides, de 5,2% de fibres et de 1% restants d’un mélange de vitamines et de minéraux.
Les régimes ont été préservés en argon Ambiance et administré quotidiennement aux souris pour éviter l’oxydation de l’AGPI-cl. Tout au long de l’étude, le contrôle du poids et de l’apparence a été effectué.
douze heures avant sacrifier une dermatite de contact a été induite dans l’une des oreilles de chaque souris. L’agent irritant utilisé était de 0,4-dinittro-1-fluorobenzen à 0,5% dans une solution d’acétone: huile d’olive 4: 1. 80 μL a été utilisé à partir de cette solution, qui ont été placés dans chaque oreille7.
Le sacrifice a été effectué au moyen d’une injection intrapéritonéale de pentothal de sodium (50 mg / kg), puis le sang a été extrait par une ponction cardiaque dans des tubes contenant de l’EDTA (60 mg / ml). Le sang a été centrifugé (3 500 tr / min) 10 minutes à 4 ° C et des aliquotes sériques ont été collectés et stockés à -80 ° C
Traitement des oreilles
Les deux oreilles de chaque souris étaient lourdes solde de précision. La moitié d’entre eux ont été introduites pendant 24 heures dans une cuisinière à 60 ° C et plus tard, leur poids sec et leur teneur en eau conséquente a été analysée. La moitié restante, a été homogénéisée indépendamment dans le tampon hatib (bromure hexadécyltriméthylammonium) à la proportion de 1/20 pour procéder à la mesure de la glutathione totale et de l’activité de la myeloperoxidase. Lesdites mesures ont été fabriquées comme décrit ci-dessus8.9.
Culture cellulaire
Les liaisons de chaque souris ont été extraites et cultivées comme décrit ci-dessus10. La moelle osseuse du fémur et le tibias de chaque souris ont été extraites par injection de milieu et cultivé, après différenciation, comme décrit ci-dessus1l.
dosage de prolifération
Les dérivés de la rate des lymphocytes ont été cultivés dans Plaques de 24 puits (2 x 106, cellules / puits) avec un moyen de 500 ul et incubé en présence ou en absence d’un (5 μg / ml) ou de LPS (50 μg / ml). Thridite tritié (Amersbam Biosciences, Heights Arlington, IL) à la concentration de 1 μCI / ml a été ajouté à chaque puits. L’incorporation de thymidine après 48 heures a été analysée comme décrit ci-dessus1L. Les mesures ont été effectuées en duplicata et étaient représentées comme la moyenne ± SD.
Production de cytokine
Les lymphocytes de la rate ont été cultivés dans des plaques à 6 puits (5 x 106 cellules / ml) avec 5 ml de milieu et ont été stimulés avec A et LPS aux mêmes concentrations utilisées dans le test de prolifération. Après 48 heures, les surnageants ont été collectés. Le sédiment a été lysé avec 1 ml de trizol pour une extraction d’ARN ultérieure. Dans le cas de macrophages, ceux-ci ont été cultivés dans des plaques de 6 puits (3 x L06 cellules / puits) et stimulées avec des LPS (100 ng / ml). Après 24 heures, le surnageant a été collecté et des sédiments cellulaires ont été lysés.
IGA, les niveaux IgE et IgG1 ont été déterminés en sérum. La concentration de l’IL-10, TNFα et IL-2 a été mesurée dans une rate surnageant et des macrophygos par ELISA après les instructions du fabricant.
analyse statistique
Le degré de statistiques de signification entre le Les groupes ont été déterminés par le test de l’étudiant T sur deux files d’attente et avec des variances égales entre les échantillons (p < 0,05). Les résultats ont été exprimés comme la moyenne ± SD.
résultats
Les animaux ont été contrôlés au cours du mois que l’expérience a duré l’observation d’un comportement et d’une apparence normaux. Il a été constaté que l’évolution de la croissance était adéquate et qu’il n’y avait pas de différence entre les deux groupes.Il n’y avait pas de type d’anomalies en ce qui concerne la quantité, la cohérence et l’apparence du tabouret, étant la teneur en eau du même similaire dans les deux groupes.
dans le but d’évaluer les effets des acides gras w -3 dans les rongeurs divers paramètres systémiques ont été étudiés qui pourraient fournir des informations sur la manière dont ils modulent le système immunitaire et clarifierent donc leurs mécanismes d’action possibles.
Les cytokines sont un groupe de molécules très divers produits par des cellules dans réponse à une grande variété de stimuli et qui sont capables de modifier en quelque sorte le comportement d’autres cellules. C’est le cas de l’IL-10, une interleup d’un type de réglementation qui médiate la réponse soi-disant Th3. Dans cette étude, le sérum, le surnageant de la culture de lymphocytes et sur le surnageant de macrophages et les niveaux dudit médiateur ont été déterminés par ELISA, a découvert une augmentation significative du groupe de souris nourries avec des acides gras W-3 dans la production d’IL -10 par macrophages (94,7 ± 57,8 VS 126,5 ± 37,6 PG / ml, p = 0,05) et lymphocytes (31,56 ± 5,51 VS 61,29 ± 29.08 pg / ml p = 0,045) étant les cellules stimulées (fig. L).
Fig. 1.-La consommation d’acides gras W-3 induit l’expression de l’IL-10. Niveaux IL-10 dans la culture de la rate surnatant Sans stimulée ou stimulée avec Concanvaline A (a).
à la suite de l’augmentation de l’IL-10, la diminution des cytokines a été observée de type TYPE, telle AS IL-2 publié par des lymphocytes (332,83 ± 197,6 VS 181.61 ± 137.1 pg / ml, p = 0,05) (Fig. 2a). La diminution des niveaux de cette interleukine a entraîné la diminution significative de la prolifération de lymphocytes B dans le groupe de souris traitées avec W-3 (7,300 ± 2,873,55 VS 4.681 ± 2,243,35 cpm, p = 0,04) déjà une réduction Dans la prolifération des lymphocytes T (7 622,6 ± 3,692,8 VS 5,048,4 ± 2,507,07 cpm, p = 0,07) pointant déjà et un éventuel effet suppresseur des acides gras W-3 sur le système immunitaire (figure 2b). Ces données sont également corrélées avec une tendance à diminuer la taille de la rate exprimée sous forme de souris mg de rate / g (5,72 ± 1,36 VS 4,7 ± 0,54, p = 0,08).
Cette diminution du nombre de lymphocytes doit avoir un impact drastique sur le sujet en raison du rôle fondamental de ces cellules dans la défense de l’organisme. Par conséquent, d’autres cytokines libérés par ces cellules pourraient être modifiées. L’effet de l’IL-10 devrait également avoir rechuté sur des macrophages afin de vérifier si une telle corrélation est survenue, certains de ces donores de supports, tels que TNFα (5,306 ± 0,51 vs 3.831 ± 0,82 ng / ml, p = 0,06) (fig. 2c).
En plus de la diminution de la réponse de type TH1, nous avons également observé que le régime riche en W-3 a exercé un effet inhibiteur de la réponse TH2 comme observé par la diminution des niveaux de plasma IgE (291, 7 ± 53,06 vs 176.04 ± 71,4 ng / ml, p = 0,06) et IgG (802.31 ± 228,1 VS 392.01 ± 191.3 μg / ml, p = 0,05) (figure 3A, B) ou IL-4 et histamine (données non représentées )) de faiblesse ou de prédisposition à l’infection. Cela peut être dû à l’augmentation de ces animaux paramètres liés à la défense naturelle de l’organisme en tant que niveaux d’IgA sérum (208,23 ± 100,3 -vs 572,26 ± 135,01 μg / ml, p = 0, 04) (figure 3C), expression de Ino et la production de Macrophages non présentés (données non représentées).
Tous ces résultats suggèrent un rôle anti-inflammatoire des acides gras G-3. Pour réaffirmer ce document, un modèle de dermatite de contact a été utilisé dans ces mêmes souris où la réponse TH1 et TH2 interviennent. Il a été réalisé dans les oreilles et l’analyse des mêmes informations contributives sur le déroulement de l’inflammation et des conséquences palpables de l’administration d’acides gras W-3.
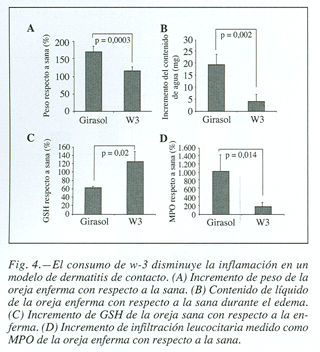
après induction de la dermatite contact, les oreilles ont été coupés et ont rapidement analysé leurs poids, observant une augmentation accrue de poids en raison de la formation d’œdème, en ce qui concerne la santé en bonne santé, dont la régie contenait uniquement de l’huile de tournesol (168,041 ± 20,16 VS 113,024 ± 11.9 exprimée en%, p = 0,0003) (Fig. 4a); Son poids sec a été déterminé en étant considéré que des valeurs similaires ont été acquises dans les deux groupes et, par conséquent, l’augmentation accrue du poids est exclusivement due à l’extravase de liquides et à la migration cellulaire vers l’orientation inflammatoire. La teneur en liquide des oreilles a été évaluée, vérifiée qu’elle était également beaucoup plus élevée dans le groupe de souris nourries aux acides gras W-6 (19,4 ± 4,5 Vs 3.94 ± 2,92 mg, p = 0,002) (figure 4b).
D’autres paramètres offrant plus de données ont été déterminés sur le concours de processus inflammatoires, en ce sens, glutathion des oreilles de souris des régimes consommés riches en acides gras W-3, il était significativement supérieur au groupe témoin (61,04 ± 4,5 VS 123,69 ± 24,2 exprimé en%, p = 0,02) (figure 4C).
Le plus grand La gravité de l’inflammation a également été corroborée avec la mesure de l’activité de la myElperoxidase. Cette enzyme est présente dans les granules de neutrophiles et catalyse la production de HCLO, constituant un marqueur sensible de l’infiltration et une inflammation de neutrophiles. Une plus grande infiltration leucocarieuse a été observée dans les oreilles enflammées de souris qui avaient consommé de l’huile de tournesol (163,19 ± 52,9 contre 1 009,74 ± 404,69 exprimé en%, p = 0,014) (figure 4D).
Tout Ces résultats indiquent que l’incorporation dans le régime alimentaire des acides gras G-3 produit un règlement sur le système immunitaire qui provoque l’individu moins prédisposé à subir des processus inflammatoires. La modification d’un grand nombre de paramètres systémiques, parmi lesquelles l’augmentation de l’IL-10 se distingue, pourrait être responsable de la protection exercée par les acides gras G-3 du régime alimentaire.
Discussion
Parmi les acides gras, ceux de la série W-3 semblent être ceux avec l’activité immunomodulante la plus élevée et dans le W-3, sont les dérivés du poisson, de l’EPA et du DHA, de manière biologique plus active4. De nombreuses expériences chez les animaux et les études d’intervention clinique indiquent que les acides gras G-3 ont des propriétés anti-inflammatoires et pourraient donc être utiles dans le traitement des maladies inflammatoires et auto-immune.
La première preuve d’importance à inclure Les acides gras G-3 dans le régime alimentaire au niveau de l’inflammation, ont émergé d’observations épidémiologiques de faible incidence de maladies auto-immunes et inflammatoires dans les Esquimements de Groenland. Peu plus tard, il y avait une période d’expansion dans la connaissance de la PUFAS en général, et du W-3 en particulier4.
On sait actuellement que les acides gras G-3 sont essentiels à la croissance normale et développement de l’individu et qui jouent un rôle clé dans la prévention et le traitement des maladies cardiovasculaires, de l’hypertension ou de l’arthrite parmi d’autres4.
Malheureusement, dans les régimes occidentaux, la consommation d’acides gras excessives est prédominate, métaboliquement différent de W-3 et avec des fonctions physiologiques généralement opposées. Ce déséquilibre contribue à l’augmentation de l’incidence des problèmes cardiovasculaires et des troubles inflammatoires4. Pour tout cela, cela devient un défi pour de nombreux scientifiques, démontrant et explication à l’effet anti-inflammatoire des acides gras W-3.
Nos résultats suggèrent que les acides gras W-3 diminuent la prolifération des lymphocytes probablement. En raison de l’effet qu’ils exercent sur la production d’IL-2 et d’autres cytokines impliquées dans la prolifération et la différenciation des lymphocytes. Ces mêmes résultats ont été trouvés par de nombreuses études sur des animaux expérimentaux12,13. Dans des études cliniques chez l’homme où les acides gras ont été inclus W-3, ces paramètres ont également été analysés, étant une diminution de la prolifération des lymphocytes, bien que dans ce cas, sans affecter les niveaux de l’IL-214. Cependant, les résultats fournis par des études chez l’homme sont contradictoires, trouvant des travaux, y compris le même groupe, où il n’est pas modifié ni même augmente la prolifération des lymphocytes. Ces différences peuvent être dues à: les doses des acides gras testés dans des études humaines ne peuvent pas être comparées aux doses élevées administrées chez les animaux; Les préparations d’huile de poisson utilisées dans différentes études varient de leurs concentrations EPA et DHA peuvent également varier les effets, et finalement la plupart des études humaines n’ont pas été suffisamment cohérentes pour prendre en compte la grande variation de certains paramètres de la réponse immunologique.
Les acides gras W-3, en plus de réduire la production de cytokine IL-2, diminue une autre série de cytokines également TH1, telles que IL-1β et TNFα produites par des macrophages. Dans les humains en bonne santé, les niveaux des deux cytokines sont également régis par les acides gras du G-317 suggérant que l’effet anti-inflammatoire pourrait être médié par la modulation exercée sur ces cytokines.
par rapport à la réponse TH2 Notre étude montre également une diminution de la même chose depuis que nous avons observé une réduction des niveaux sériques d’histamine, d’IG et de LGG1, ainsi que des cytokines telles que IL-4. Il existe diverses preuves épidémiologiques qui soutiennent le rôle protecteur des acides gras W-3 dans les maladies allergiques.Cependant, des études utilisant des suppléments de poisson dans l’asthme révèlent un impact clinique rare malgré des changements biochimiques importants qui se produisent18. Même dans ce cas, de nouvelles études sont menées dans l’asthme et d’autres atopies afin d’améliorer le cours de la maladie.
Divers auteurs défendent un autre type d’acides gras autres que W-3 pour penser que je pouvais obtenir là à un état d’immunosuppression qui a laissé l’individu non protégé et a commis son état de santé19. Cependant, nos résultats suggèrent que les acides gras G-3 sont en mesure de renforcer certaines défenses de l’organisme, telles que l’immunoglobuline A, dont l’augmentation peut également être renforcée par l’augmentation de l’IL-10 qui induit le changement d’isotycle des immunoglobulines. Dans cette étude, l’augmentation de l’expression des inos dans des macrophages qui impliquerait une aide contre une attaque externe est également observée.
À notre avis, le résultat le plus pertinent et le plus roman de cette étude est l’augmentation de la IL-10 Cytokine réglementaire, qui pourrait être considérée comme responsable de l’effet anti-inflammatoire des acides gras G-3, ainsi que antiallergique, car le pouvoir immunosuppresseur de ladite cytokine pourrait diminuer les réponses à la fois TH1 et TH2.
Diverses études, telles que celles effectuées sur des modèles de pancréatite expérimentale20 où la nutrition parentérale a été appliquée avec de l’huile de poisson, coïncidant dans lesquelles les acides W-3 induisent une augmentation des niveaux IL-10. Cependant, d’autres auteurs n’observent pas les différences21 ou même percevoir une diminution de l’IL-1022 dans d’autres modèles. Il serait souhaitable de s’approfondir de ce fait à tenter de spécifier le rôle de l’IL-10 dans le mécanisme d’action des acides gras du G-3, raison pour laquelle les études avec assassinement pour l’IL-10 devraient être effectuées Dans nos laboratoires.
Le modèle de dermatite de contact utilisé dans cette étude permet d’observation à l’œil nu de l’effet anti-inflammatoire de l’alimentation enrichie dans les acides gras G-3. Les valeurs de poids des oreilles et la teneur en eau du même reflètent déjà l’effet d’amieflammatoire clair de l’huile de poisson. Que la teneur en poids et en eau des oreilles enflammées de souris nourries avec W-3 était mineure était indicative que, dans ledit groupe, la vasodilatation avait été moins drastique et donc le moindre œdème. Les études japonaises confirment ces effets en mode similaire, attribuant dans ce cas l’effet à la plus grande puissance du DHA7. Une étude similaire, utilisant des esters phorbol (TPA) en tant qu’agent irritant, indique également une puissance anti-inflammatoire, dans ce cas, de la DHA et de l’EPA23.
mesure MPO montre une infiltration de leucocytes inférieure avec le traitement de Acides gras W-3. La migration des leucocytes de la microvasculature au centre de la blessure où ils s’accumulent, est une étape clé caractéristique de l’inflammation aiguë23,7. Les acides gras du G-3 semblent avoir un rôle important à ce niveau car dans ledit groupe, le phénomène est à peine perçu.
Les radicaux libres associés à une inflammation entraînent un stress oxydatif, impliqué dans l’aggravation de nombreuses maladies. Le glutathion est un tresptide de haute capacité antioxydante auquel l’organisme a recours à la neutralisation des radicaux libres. Dans les oreilles de souris où la dermatite de contact a été induite des niveaux de glutathion diminuent de manière significative lors de la gestion du stress oxydatif généré. Toutefois, dans le groupe W-3, cette diminution est réduite ou compensée depuis l’étant plus faible dommage, son effet antioxydant n’est pas si nécessaire. Ces résultats pourraient suggérer que les acides gras G-3 augmentent les niveaux de glutathion, mais lors de l’analyse de la teneur en glutathion dans le foie de tous les rongeurs, aucune différence n’a été observée. Cependant, il existe des études relatives à la consommation d’acides gras W-3 avec une augmentation des défenses antioxydantes24,25.
Nous pouvons conclure les attribuer aux acides gras W-3 un puissant papier anti-inflammatoire où la L’augmentation de l’IL-10 semble être le facteur de déclenchement d’une série de modifications de la réponse immunologique conduisant à cet effet. La diminution de la prolifération des lymphocytes et la réponse du type TH1 et TH2 sont certaines de ces modifications. L’étude de l’effet des acides W-3 au niveau des voies de transduction de signal est essentielle pour terminer une explication à la grande activité de ces composés, non seulement au niveau de l’inflammation, mais également au niveau d’allergie.
Références
1. Kelley D: modulation de l’immunité humaine et des réponses inflammatoires par des acides gras alimentaires. Nutrition 2001, 17 (7/8): 669-673.
2. Calder PC: acides gras polyinsaturadés N-3 et inflammation: de la biologie moléculaire à la clinique. LIPIDES 2003, 38: 323-352.
3.Harbige LS: acides gras, réponse immunitaire et auto-immunité: une question de l’essentialité N-6 et de l’équilibre entre N-6 et N-3. LIPIDES 2003, 38: 323-341.
4. SIMOPOULOS AP: acides gras oméga-3 dans l’inflammation et les maladies autoinnummumes. J Am Coll Nut 2002,21 No.6: 495-505.
6. Yaqoob P: lipides et réponse immunitaire: du mécanisme moléculaire aux applications cliniques. Cur d’Opin Nutte Métab Care 2003, 6: 133-150.
7. Tomobe Yi, Morizawa K, Tsuchida M, Hibino H, Nakano Y, Tanaka Y: Acide diététique Docosahexaénoïque supprime l’inflammation et les immunoresponses dans la réaction d’hypersensibilité à la souris. LIPIDES 2000, 35, no. 1: 61-69.
8. Anderson Me: Détermination du glutathion et du glutathion disulfure dans des échantillons biologiques. Enzymol de méthamphétamine 1985, 113: 548-555.
9. Krawisz Ye, Sharon P, STENSON WF: test quantitatif pour une inflammation intestinale aiguë basée sur l’activité de la myeloperoxidase. Évaluation de l’inflammation chez le rat et les modèles de hamster. Gastroenterology 1984, 87: 1344-1350.
10. Kelley DS, Warren JM, Simon VA, Bartolini G, Mackey Be, Erickson KL: Effets similaires de C9, T11-CLA et T10, CL2CLA sur les fonctions de cellules immunes chez la souris. LIPIDES 2002, 37: 725-728.
11. Xaus J, Cardó m, Vellador AF, Soler C. Lloveras J, Celada A: Interféron Gamma induit l’expression de P21WAF1 et arrête le cycle de cellules de macrophage empêchant l’induction de l’apoptose. Immunité 1990, 2: 103-113.
12. Peterson LD, Thies F, Sanderson P, Newsholme Ea, Calder PC: Les faibles niveaux d’acides eicosapentaénoïque et docosahexaénoïque imitent l’effet de l’huile de poisson sur les lymphocytes de rat. LIFE SCI 1998, 62 (24): 2209-2017.
13. Peterson Ld, Jeffery Nm, Thies F, Sanderson P, Newsholme EA Y Calder PC: Les acides eicosapentaénoïque et docosahexaénoïque Alter Rat Scheen Les acides gras de leucocyte Composition et prostaglandine E2 Production mais ont des effets différents sur la fonction de lymphocytes et l’immunité médiatisée de la cellule cellulaire. LIPIDES 1998, 33 (2): 171-180.
14. Thies F, Neveu-von-Caron G, Powell Jr, Yaqoob, P, Newsholme Ea, Calder PC: La supplémentation alimentaire avec l’acide gamma-linolénique ou l’huile de poisson diminue la prolifération de lymphocytes chez l’homme plus âgé sain. J NUL 2001, 131 (7): 1918-1927.
15. Kew S, Benerjee T, Minihane am, Finnegan Ye, Muggli R, Albers R, Williams CM, Calder PC. Manque d’effet des aliments enrichis d’acides gras de N-3 dérivés de plante-ormarine sur les fonctions immunitaires humaines. Am J Clin Clin Nut 2003, 77 (5): 1287-1295.
16. Trebble TM Y Cols .: Protamblandin E2 Production et T-Cellule T Après la supplémentation de l’huile de poisson: Réponse à la costuplement antioxydant. Am J Clin Clin Nut 2003, 78 (3): 376-382.
17. Endres S Y Cols.: Effet de la complémentation alimentaire avec les acides gras N-3 sur la synthèse du facteur de nécrose interleukine-1 et de la nécrose tumorale par des cellules mononucléées. N ENGL J MED 1989, 320 (5): 265-271.
18. Calder PC, Yaqoob P, Thies F, Wallace Fa Y Miles EA: Acides gras et fonctions de lymphocytes. Brit J Nutal 2002, 87 Suppl. 1, S31-S48.
19. Puertollano Ma, de Pablo M, Álvarez de Cienfuegos G: Les propriétés anti-oxydantes de la N-acétyl-kyscine n’améliorent pas la résistance immunitaire des souris nourries aux lipides alimentaires de la Listeria monocytogenes. Clin NIP, 2003, 22 (3): 313-319.
20. FOITZIK T, EIBL G, Schneider P, Wenger Fa, Jacobi CA, BUHR HJ: La supplémentation en acides gras oméga-3 augmente les cytokines anti-inflammatoires et atténue les séquelles de maladie systémique dans la pancréatite expérimentale. PARDER ENTREAL ELL, 2002, 26 (6): 351-356.
21. Wallace Fa, Miles Ea, Calder PC: Comparaison des effets de l’huile de lin et différentes doses d’huile de poisson sur la fonction de cellule mononucléée chez les sous-réjets humains sains. BR J NUT 2003, 89 (5): 679-681.
22. Hayashi Y Cols.: Effets de l’émulsion intraveineuse oméga-3 et de la graisse oméga-6 sur la production de cytokine et la sensibilité de l’hiperficie de type retardée dans les rats brûlés recevant une nutrition parentérale totale. J POUERER ENTERAL ELLE 1998, 22 (6): 363-367.
23. Raerlerstorff D, Panteze M, Bachmann H, Moser U: Propriétés anti-inflammatoires des acides docosahexaénoïque et d’eicosupentaénoïque dans une inflammation d’oreille de souris induite par l’ester phorbol. Int Arch Allergy Immunol 1996, 111 (3): 284-290.
24. Jolly CA, Muthukumar A, Avula CP, Troyer D, Fernandes G: La durée de vie est prodigée dans une émission auto-immune restreinte de nourriture (NZBXNZW) F (L) Les souris ont nourri un régime enrichi d’acides gras (N-3). J NUL 2001, 131 (10): 2753-60.
25. Demoz A, Willunsen N Y Berge RK: L’acide eicosapentaénoïque à la dose hypotriglycéridémique améliore la défense antioxydante hépatique chez les souris. LIPIDES 1992, 27 (12): 968-971.