alimenti funzionali
l’espressione di IL-10 interviene nel regolamento della risposta infiammatoria di Omega 3 acidi grassi
s. Sierra, F. Lara-Villoslada, M. Olivares, J. Jiménez, J. Boza e J. Xaus
Dipartimento di immunologia e studi preclinici. Puleva Biotech. Granada.
|
Riepilogo Target o sfondo: acidi polinosaturati I grassi sono importanti per l’organismo umano dovuto al suo coinvolgimento in numerose funzioni biologiche. Le diete occidentali sono caratterizzate da essere eccessivamente ricche di acidi grassi W-6 e poveri di acidi grassi W-3. (Nut of Hosp 2004, 19: 376-382) Parole chiave: acidi grassi polune. Infiammazione. Organismo. |
L’espressione IL-10 è indispensabile nel regolamento Della risposta immunitaria di omega 3 acidi grassi astratto introduzione: gli acidi grassi polinsararati svolgono un ruolo chiave in un numero enorme di funzioni biologiche. Le diete occidentali sono molto ricche di acidi grassi W-6. Tuttavia il contenuto degli acidi grassi W-3 non è adatto a quelle diete, spremire della loro importanza nel normale sviluppo del corpo umano e del regolamento della risposta immunitaria. (Nut-Hosp 2004, 19: 376-382) Parole chiave: Acidi grassi polinsaturi. Antinfiammatorio Organismo. |
corrispondenza: J. Xaus.
Dipartimento di immunologia e studi preclinici.
puléva biotech.
Road de Purchel, 66. 18004 Granada.
E-mail: [email protected]
ricevuto: 21-I-2004. Accettato: 13-II-2004.
Introduzione
Il sistema immunitario è coinvolto nella difesa dell’organismo contro agenti stranieri, cellule tumorali e aggressioni esterne. Un alto numero di organi, cellule, tessuti e molecole interagiscono per svolgere questa missione.
Lo sviluppo, la manutenzione e il funzionamento ottimale del sistema immunitario dipendono dalla dieta tra gli altri fattori. Tra i nutrienti che più influenzano il sistema immunitario evidenzia il grasso la cui carenza o il contributo squilibrato nonché la qualità di compromette la stabilità del sistema immunitario1.
Il rapporto tra acidi grassi e infiammazione è dovuto principalmente al Fosfolipide dei componenti delle cellule del sistema immunitario. Gli acidi grassi incorporati nella membrana di loro possono influenzare le funzioni di esso e quindi il funzionamento cellulare. I fosfolipidi della membrana sono responsabili della fluidità, che influenzerà i diversi componenti ancorati in esso. Diversi segnali intracellulari partono da questi fosfolipidi e dalla loro composizione, quindi influenzano diversi percorsi di trasduzione del segnale. Inoltre, gli acidi grassi sintetizzati dai fosfolipidi sono substrati per la sintesi di un gruppo di mediatori, eicosanoidi, marcata importanza biologica2.
Attualmente, l’interesse nello studio degli acidi grassi è in aumento e l’influenza di loro è importante sulla risposta immunologica. Questo interesse si è concentrato sullo studio di acidi grassi polinsaturati a catena lunga (AGPI-CL), provenienti dalle famiglie W-6 e W-3, i cui precursori, acidi linoliici e α-linolenici sono essenziali, dover essere ingerito con il Dieta, così come si consiglia inoltre di includere alcuni dei suoi derivati, come l’acido eicosapentaenoico (EPA), Docosahexaenoic (DHA) e Arachidonic (AA), da quando attraverso i percorsi metabolici, con l’allungamento dei suoi precursori, i livelli appropriati sono Non raggiunto3.
In individui che consumano diete occidentali, le cellule infiammatorie contengono elevate proporzioni di AGPI-CL della serie W-6, in particolare AA e bassa proporzioni, tuttavia, dall’EPA del W-34 serie. Il ruolo principale dell’AA è quello di agire come un substrato per la sintesi di mediatori bioattivi del tipo eicosanoide. Questi sono coinvolti in modulare la durata e l’intensità della risposta infiammatoria. I derivati AA, come TXA2 e LTB4, sono agenti potenti con un marcato effetto infamato. Il TXA2, ha un vasocostrittore e un effetto proaprenico mentre il LTB4 è un potente agente chemiotattico5. Inoltre, il PGE2 induce la febbre, aumenta la permeabilità vascolare e la vasodilatazione, mentre la produzione di cellule IGE b.
l’aumento del consumo di acidi grassi W-3 porta a una diminuzione in AA nelle membrane di Le cellule infiammatorie riducono così la loro capacità di sintetizzare dal Cox e dagli eicosanoidi derivati dall’AA. Oltre ad inibire il metabolismo AA, l’EPA inibisce l’ossigenazione AA da parte di Cox e sopprime l’induzione di alcune citochine che agiscono sull’espressione genica di Cox-2 e 5-Lox. È anche in grado di agire come un substrato di COX e 5-LOX, dando origine ad eicosanoidi considerati biologicamente meno attivi5.
Tutto ciò porta all’idea che l’olio di pesce ha proprietà antinfiammatorie. Il consumo in individui sani di una proporzione adeguata degli acidi grassi G-6 e W-3 porterebbe a una migliore regolamentazione della risposta immunologica, e il conseguente aumento della proporzione di acidi grassi W-3 sarebbe vantaggioso per i soggetti che soffre di malattie autoimmuni o infiammatorie a causa dell’effetto antinfiammatorio di loro1.
Non solo questi meccanismi sono coinvolti nell’effetto antinfiammatorio, ma il loro coinvolgimento del percorso di segnalazione è attualmente conosciuto come NFKB, MAPK o PPAR e LXR6. L’azione attraverso questi due recettori apre un nuovo campo di ricerca e una gamma maggiore di applicazioni degli acidi grassi G-3 dovuta alla posizione e alla funzione di esse.
Lo studio attuale è quello di dimostrare l’anti- Effetto infiammatorio degli acidi grassi G-3 e dei meccanismi coinvolti, misurando parametri immunologici che consentono di controllare il suo effetto a livello sistemico e analizzare il corso di un’infiammazione indotta per determinare il suo livello locale.
Materiale e metodi
Reagenti e anniversanti
Concanvalin A (con A) e Lipopolisaccaride (LPS) sono stati forniti da Sigma Chemical Co (St Louis, Mo). Gli anticorpi forniti da Bethyl (Montgomery TX) sono stati utilizzati per misurare le immunoglobuline e nel caso delle citochine da Bioosource (Camarillo, CA). Perossidasi ravanello, bromuro di esadeciltrimetilmonium, o-dianysidina cloridrato e perossido di idrogeno per la misurazione dell’attività di milleoperoxidasi, nonché l’acido tricloroacetico, è stato anche acquisito β-nadpa, 5,5-ditio-bis (nitrobenzoico a 2 acidi) e glutatione sono stati acquisiti anche in Sigma . Glutatithione Reductase è stato fornito da Roche.L’acqua deionizzata purificata è stata utilizzata con un sistema Milli-Q (Bedford, MA).
Animali e diete
Balb / c topi maschili per svezzare tre settimane di età sono state fornite dall’università di Granada. Sono stati mantenuti per tutto l’esperimento a una temperatura controllata di 22 ° C e sotto un ciclo di luce di 12 ore. L’esperimento globale è stato effettuato 3 volte con risultati simili.
Tutti gli studi sugli animali che sono stati effettuati sono stati effettuati secondo gli standard etici animali impostati nei criteri di Helsinki.
L’esperimento Consiste di 2 gruppi di topi (N = 10) alimentati per 28 giorni con diete di base uguali e la cui differenza risiede nel tipo di grasso incluso. Entrambi contengono il 5% di grasso della dieta totale. Questo grasso del 5% è costituito da olio di girasole nel caso del gruppo utilizzato come controllo. Nel caso della dieta W-3, il 12% del contenuto di grassi totale è raffinato olio di pesce, mentre il resto del grasso è lo stesso olio di girasole dal gruppo precedente. Entrambe le diete sono state elaborate da specifiche run-up seguendo le raccomandazioni dell’American Veterinary Academy for Animal Nutrition. In questo modo sono state preparate le diete semi-sintetiche, incluso il 5% di grassi, la proteina del 21,3%, i carboidrati del 67,5%, il 5,2% di fibre e il restante 1% di una miscela di vitamine e minerali.
Diete sono state conservate in Argon Atmosfera e somministrata quotidianamente ai topi per evitare l’ossidazione dell’AGPI-Cl. Durante tutto lo studio, il controllo del peso e dell’aspetto è stato effettuato.
dodici ore prima del sacrificio Una dermatite da contatto è stata indotta in una delle orecchie di ogni mouse. L’agente irritante utilizzato era 0,4-Dinitro-1-fluorobenzen a 0,5% in una soluzione acetone: olio d’oliva 4: 1. 80 μL è stato utilizzato da questa soluzione, che sono stati collocati in ciascun orecchio7.
il sacrificio è stato eseguito mediante un’iniezione intraperitoneale di sodio Pentothal (50 mg / kg) e poi il sangue è stato estratto dalla foratura cardiaca Nelle provette contenenti EDTA (60 mg / ml). Il sangue era centrifugato (3.500 giri / min) 10 minuti a 4 ° C e aliquota sierica sono stati raccolti e conservati a -80 ° C.
Elaborazione delle orecchie
Entrambe le orecchie di ogni topo erano pesanti in Saldo di precisione. La metà di loro è stata introdotta per 24 ore in una stufa a 60 ° C e in seguito il loro peso secco e il loro conseguente contenuto di acqua è stato analizzato. La metà rimanente, è stata omogeneizzata in modo indipendente nel buffer Hitab (Bromuro esadecyltrimetilammonio) alla proporzione di 1/20 per procedere con la misurazione dell’attività totale glutatione e mieloperossidasi. Tali misurazioni sono state realizzate come descritto sopra8.9.
Cultura cellulare
Le obbligazioni di ciascun mouse sono state estratte e coltivate come descritto sopra10. Il midollo osseo del femore e le tibie di ogni topo sono stati estratti dall’iniezione di media e coltivati, dopo la differenziazione, come descritto sopra1L.
test di proliferazione
i linfociti derivati milzan sono stati coltivati Piatti da 24 pozzetti (2 x 106, celle / pozzetti) con 500 UL Medium e incubati in presenza o assenza di A (5 μg / ml) o LPS (50 μg / ml). Timidite triziata (Bioscienze Amersbam, Arlington Heights, IL) alla concentrazione di 1 μCI / ML è stata aggiunta a ciascun pozzetto. L’incorporazione della timidina dopo 48 ore è stata analizzata come descritto sopra1L. Le misure sono state eseguite in duplicate e sono state rappresentate come media ± SD.
Produzione di citochine
I linfociti della milza sono stati coltivati in piatti 6 pozzetti (5 x 106 celle / ml) con 5 ml di medium e sono stati stimolati con A e LPS alle stesse concentrazioni utilizzate nel saggio di proliferazione. Dopo 48 ore sono stati raccolti i supernatanti. Il sedimento è stato lisato con 1 ml di trizol per la successiva estrazione dell’RNA. Nel caso di macrofagi, questi sono stati coltivati in piatti a 6 pozzetti (celle di 3 x L06 / bene) e stimolate con LPS (100 ng / ml). Dopo 24 ore il Supernatante è stato raccolto e i sedimenti cellulari sono stati pelosi.
IGA, livelli IGE e IgG1, sono stati determinati nel siero. La concentrazione di IL-10, TNFα e IL-2 è stata misurata in milza supernatant e macrofotografia da ELISA seguendo le istruzioni del produttore.
Analisi statistica
Il grado di significatività tra il I gruppi sono stati determinati dal test T-Student su due code e con varianze uguali tra i campioni (P < 0,05). I risultati sono stati espressi come media ± SD.
Risultati
Gli animali sono stati controllati durante il mese in cui l’esperimento è durato osservando un comportamento e un aspetto normale. Fu trovato che l’evoluzione nella crescita era adeguata e che non c’erano differenze tra i due gruppi.Non c’era nessun tipo di anomalie rispetto alla quantità, consistenza e aspetto dello sgabello, essendo il contenuto d’acqua dello stesso simile in entrambi i gruppi.
con lo scopo di valutare gli effetti degli acidi gras w -3 In Roditori sono stati studiati vari parametri sistemici che potrebbero fornire informazioni su come modulano il sistema immunitario e pertanto, chiariscono i loro possibili meccanismi di azione.
Le citochine sono un gruppo molto diverso di molecole prodotte da celle in Risposta a un’ampia varietà di stimoli e che sono in qualche modo in qualche modo alterando il comportamento di altre cellule. Questo è il caso di IL-10, un’interleucca di un tipo normativo che media la cosiddetta risposta TH3. In questo studio, il siero, il supernatante della coltivazione dei linfociti e del soprannatante di macrofagi e dei livelli di detto mediatore sono stati determinati da Elisa, ha scoperto un aumento significativo del gruppo di topi nutriti con acidi grassi W-3 nella produzione di IL -10 di Macrofagi (94,7 ± 57,8 VS 126,5 ± 37.6 pg / ml, p = 0,05) e linfociti (31,56 ± 5,51 VS 61,29 ± 29.08 pg / ml P = 0,045) Essendo le celle stimolate (fig. l).

Fig. 1. – Il consumo di acidi grassi W-3 induce l’espressione di IL-10. LIVELLI DI IL-10 IN PLEENURE CULTURA SUPERNATANTE
Stimolato o stimolato con Concanvalin A (A).
A seguito dell’aumento di IL-10, è stata osservata la diminuzione delle citochine, tale Come il-2 rilasciato da linfociti (332,83 ± 197.6 vs 181.61 ± 137.6 pg / ml, p = 0,05) (figura 2a). La diminuzione dei livelli di questa interleucca ha portato alla significativa diminuzione della proliferazione dei linfociti B nel gruppo di topi trattati con W-3 (7.300 ± 2,873.55 vs 4.681 ± 2.243.35 cpm, p = 0,04) già una riduzione Nella proliferazione dei linfociti T (7.622,6 ± 3.692.8 vs 5,048,4 ± 2,507.07 cpm, p = 0,07) già indicando e un possibile effetto soppressore degli acidi grassi W-3 sul sistema immunitario (figura 2b). Questi dati sono anche correlati con la tendenza a diminuire le dimensioni della milza espressa come mg milza / g mouse (5.72 ± 1,36 vs 4.7 ± 0,54, p = 0,08).

Questa diminuzione del numero di linfociti deve avere un impatto drastico sull’argomento dovuto al ruolo fondamentale di Queste cellule nella difesa dell’organismo. Pertanto, altre citochine rilasciate da queste cellule potrebbero essere modificate. Anche l’effetto di IL-10 dovrebbe essere ricapito su macrofagi per verificare se tale correlazione si è verificata, alcune di queste donazioni multimediali, come TNFα (5,306 ± 0,51 contro 3.831 ± 0,82 ng / ml, P = 0,06) (fig. 2c).
Oltre alla diminuzione della risposta del tipo TH1, abbiamo anche osservato che la dieta ricca di W-3 ha esercitato un effetto inibitorio della risposta TH2 osservata dalla diminuzione dei livelli di plasma IGE (291, 7 ± 53,06 vs 176.04 ± 71,4 ng / ml, p = 0,06) e IgG (802.31 ± 228,1 VS 392.01 ± 191.3 μg / ml, p = 0,05) (figura 3a, b) o IL-4 e istamina (dati non mostrati ).

Nonostante la diminuzione diffusa nella risposta immunologica, i topi alimentati da W-3 non hanno sperimentato i sintomi di debolezza o predisposizione alle infezioni. Ciò può essere dovuto all’aumento di questi animali parametri relativi alla difesa naturale dell’organismo come livelli sierici IGA (208,23 ± 100.3 -VS 572,26 ± 135,01 μg / ml, p = 0, 04) (figura 3C), espressione di Ino e la produzione di non da macrofagi (dati non mostrati).
Tutti questi risultati suggeriscono un ruolo anti-infiammatorio degli acidi grassi G-3. Per riaffermare questo documento, un modello di dermatite da contatto è stato utilizzato in questi stessi topi dove interviene sia la risposta TH1 che TH2. È stato effettuato nelle orecchie e l’analisi delle stesse informazioni contribuiscono nel corso dell’infiammazione e delle conseguenze palpabili della somministrazione di acidi grassi W-3.
Dopo l’induzione del contatto della dermatite, le orecchie sono stati tagliati e analizzati rapidamente i loro pesi, osservando un maggiore aumento del peso dovuto alla formazione di edema, rispetto alla salute, nei topi le cui diete contenevano solo olio di girasole (168,041 ± 20,16 vs 113.024 ± 11.9 espresso in%, P = 0.0003) (Figura 4a); Il suo peso secco è stato determinato essendo visto che sono stati acquisiti valori simili in entrambi i gruppi e, pertanto, l’aumento dell’aumento del peso, è esclusivamente dovuto allo stravaso di liquidi e migrazione cellulare verso la messa a fuoco infiammatoria. Il contenuto liquido delle orecchie è stato valutato, essendo controllato che è stato anche molto più alto nel gruppo di topi nutriti con acidi grassi W-6 (19,4 ± 4,5 vs 3.94 ± 2,92 mg, p = 0,002) (figura 4b).
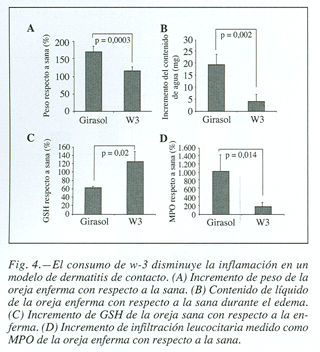
altri parametri che forniscono più dati sono stati determinati sul concorso infiammatorio del processo, in questo senso, glutatione dalle orecchie dei topi Quella dieta consumata ricca di acidi grassi W-3, era significativamente superiore al gruppo di controllo (61,04 ± 4,5 VS 123.69 ± 24.2 espresso in%, p = 0,02) (figura 4C).
Maggiore La gravità dell’infiammazione è stata anche corroborata con la misurazione dell’attività mielperossidasi. Questo enzima è presente nei granuli dei neutrofili e catalizza la produzione di HCLO, costituendo un marker sensibile dell’infiltrazione e dell’infiammazione del neutrofilo. Una maggiore infiltrazione leucocarosa è stata osservata nelle orecchie infiammate di topi infiammati che avevano consumato olio di girasole (163,19 ± 52,9 contro 1,009,74 ± 404.69 espresso in%, p = 0,014) (figura 4D).
Tutto Questi risultati indicano che l’incorporazione nella dieta degli acidi grassi G-3 produce un regolamento del sistema immunitario che causa l’individuo meno predisposto a subire processi infiammatori. La modifica di un gran numero di parametri sistemici, tra cui si distingue l’aumento di IL-10, potrebbe essere responsabile per la protezione esercitata dagli acidi grassi G-3 della dieta.
Discussione
>
Tra gli acidi grassi, quelli della serie W-3 sembrano essere quelli con la più alta attività immunomodulante e all’interno del W-3, sono i derivati del pesce, dell’EPA e del DHA, biologicamente più attivo4. Numerosi esperimenti negli animali e negli studi d’intervento clinici indicano che gli acidi grassi G-3 hanno proprietà antinfiammatorie e quindi potrebbero essere utili nel trattamento delle malattie infiammatorie e dell’autoimmune.
La prima evidenza di importanza da includere Gli acidi grassi G-3 nella dieta a livello di infiammazione sono emersi da osservazioni epidemiologiche della bassa incidenza delle malattie autoimmuni e infiammatorie nell’eschimos de Groenlandia. Poco più tardi c’era un periodo di espansione nella conoscenza dei Pufas in generale, e del W-3 in particolare4.
Attualmente è noto che gli acidi grassi G-3 sono essenziali per la normale crescita e Sviluppo dell’individuo e che svolge un ruolo chiave nella prevenzione e nel trattamento delle malattie cardiovascolari, dell’ipertensione o dell’artrite tra gli altri4.
Sfortunatamente, nelle diete occidentali il consumo di acidi grassi in eccesso W-6 predominati, metabolicamente diverso da W-3 e con funzioni fisiologiche generalmente opposte. Questo squilibrio contribuisce all’aumento dell’incidenza dei problemi cardiovascolari e dei disturbi infiammatori44. Per tutto questo, diventa una sfida per numerosi scienziati, dimostrando e spiegazione all’effetto anti-infiammatorio degli acidi grassi W-3.
I nostri risultati suggeriscono che gli acidi grassi W-3 riducono probabilmente la proliferazione dei linfociti A causa dell’effetto esercitano sulla produzione di IL-2 e altre citochine coinvolte nella differenziazione di proliferazione e linfociti. Questi stessi risultati sono stati trovati da numerosi studi in animali sperimentali12,13. Negli studi clinici negli esseri umani in cui sono stati inclusi gli acidi grassi W-3, anche questi parametri sono stati analizzati, essendo una diminuzione della proliferazione dei linfociti, anche se in questo caso senza influire sui livelli di IL-214. Tuttavia, i risultati forniti dagli studi negli esseri umani sono in conflitto, trovando alcuni lavori, compreso lo stesso gruppo, dove non è modificato o addirittura aumenta la proliferazione dei linfociti. Queste differenze possono essere dovute a: le dosi di acidi grassi testate negli studi umani non possono essere confrontate con le alte dosi somministrate negli animali; I preparati di pesciosi che vengono utilizzati in diversi studi variano che le loro concentrazioni EPA e DHA possono anche variare gli effetti, e infine la maggior parte degli studi umani non sono stati abbastanza coerenti da tenere conto della grande variazione in alcuni parametri della risposta immunologica.
Acidi grassi W-3, oltre a ridurre la produzione di cytochine IL-2, diminuisce un’altra serie di citochine anche TH1, come IL-1β e TNFA prodotto da macrofagi. Negli umani sani, i livelli di entrambe le citochine sono regolati anche dagli acidi grassi G-317 che suggeriscono che l’effetto antinfiammatorio potrebbe essere mediato dalla modulazione esercitata su queste citochine.
Rispetto alla risposta TH2 , il nostro studio dimostra anche una diminuzione degli stessi poiché abbiamo osservato una riduzione dei livelli sierici di istamina, IG e LGG1, così come citochine come IL-4. Esistono varie evidenze epidemiologiche che supportano il ruolo protettivo degli acidi grassi W-3 nelle malattie allergiche.Tuttavia, gli studi che utilizzano integratori a pesci in asma rivelano un impatto clinico scarso nonostante i significativi cambiamenti biochimici che si verificano18. Anche così, nuovi studi vengono eseguiti in asma e in altri atopia per migliorare il corso della malattia.
Vari autori difendono un altro tipo di acidi grassi diversi da W-3 per pensare che potessi ottenere lì a uno stato di immunosoppressione che ha lasciato l’individuo non protetto e ha commesso il suo stato di salute19. Tuttavia, i nostri risultati suggeriscono che gli acidi grassi G-3 sono in grado di rafforzare alcune difese dell’organismo, come l’immunoglobulina A, il cui aumento può anche essere migliorato dall’aumento di IL-10 che induce il cambiamento dell’isotipo delle immunoglobuline. In questo studio, è anche osservato l’aumento dell’aumento dell’espressione di INOS in macrofagi che implicherebbe un aiuto contro un attacco esterno.
A nostro avviso, il risultato più rilevante e romanzo di questo studio è l’aumento del Cytochine normativa IL-10, che potrebbe essere considerata responsabile dell’effetto antinfiammatorio degli acidi grassi G-3, nonché antiallergici, poiché il potere immunosoppressivo di detta citochina potrebbe ridurre le risposte sia TH1 e TH2.
Vari studi, come quelli eseguiti su modelli di pancreatite sperimentale20 in cui la nutrizione parenterale è stata applicata con olio di pesce, coincidendo in cui gli acidi W-3 inducono un aumento dei livelli di IL-10. Tuttavia, altri autori non osservano le differenze21 o nemmeno percepiscono una diminuzione del 1022 in altri modelli. Sarebbe consigliabile approfondire in questo fatto per cercare di specificare il ruolo di IL-10 nel meccanismo d’azione degli acidi grassi G-3, motivo per cui gli studi con Knock-out per IL-10 dovrebbero essere eseguiti Nei nostri laboratori.
Il modello di dermatite del contatto utilizzato in questo studio, consente l’osservazione all’occhio nudo dell’effetto antinfiammatorio della dieta arricchita in acidi grassi G-3. I valori di peso delle orecchie e il contenuto di acqua dello stesso riflettono già il chiaro effetto amiinfiammatorio dell’olio di pesce. Che il peso e il contenuto di acqua delle orecchie infiammati dei topi nutriti con W-3 erano minore era indicativo che in detto gruppo la vasodilazione era stata meno drastica e quindi il minimo edema. Gli studi giapponesi confermano questi effetti in una modalità simile, attribuendo in tal caso l’effetto al maggiore potere del DHA7. Uno studio simile, utilizzando gli esteri di Phorbol (TPA) come agente irritante, indica anche il potere antinfiammatorio, in questo caso, da DHA ed EPA23.
MPO Misura mostra un’infiltrazione di leucociti inferiori con il trattamento di Acidi grassi W-3. La migrazione dei leucociti dalla microvascolatura al focus del fulcro della ferita in cui si accumulano, è una caratteristica di fase chiave di infiammazione acuta23,7. Gli acidi grassi G-3 sembrano avere un ruolo importante a questo livello poiché in detto gruppo il fenomeno è a malapena percepito.
I radicali liberi associati all’infiammazione portano allo stress ossidativo, coinvolto nell’aggravamento di numerose malattie. Glutatione è un tripultide di alta capacità antiossidante a cui l’organismo residente per la neutralizzazione dei radicali liberi. Nelle orecchie dei topi in cui le dermatite del contatto sono state indotte i livelli di glutatione diminuiscono significativamente quando si tratta dello stress ossidativo generato. Tuttavia, nel gruppo W-3 questa diminuzione è ridotta o compensata da quando è il danno inferiore, il suo effetto antiossidante non è così richiesto. Questi risultati potrebbero suggerire che gli acidi grassi G-3 aumentano i livelli di glutatione, ma, quando analizzando il contenuto di glutatione nel fegato di tutti i roditori non sono state osservate differenze. Tuttavia, ci sono studi che si riferiscono al consumo di acidi grassi W-3 con un aumento delle difese antiossidanti24,25.
Possiamo concludere attribuendoli a acidi grassi W-3 un potente carta anti-infiammatoria dove il L’aumento delL-10 sembra essere il fattore di innesco di una serie di modifiche nella risposta immunologica che porta a questo effetto. La diminuzione della proliferazione dei linfociti e della risposta di tipo TH1 e TH2 sono alcune di queste modifiche. Lo studio dell’effetto degli acidi W-3 al livello dei percorsi di trasduzione del segnale è essenziale per finire il dare spiegazione alla grande attività di questi composti, non solo a livello di infiammazione ma anche a livello di allergia.
Riferimenti
1. Kelley D: modulazione delle risposte immune e infiammatorie umane da acidi grassi dietetici. Nutrizione 2001, 17 (7/8): 669-673.
2. Calder PC: N-3 Acidi grassi poliunsaturarati e infiammazione: dalla biologia molecolare alla clinica. Lipidi 2003, 38: 323-352.
3.Harbige Ls: acidi grassi, risposta immunitaria e autoimmunità: una domanda di essenziale N-6 e l’equilibrio tra N-6 e N-3. Lipidi 2003, 38: 323-341.
4. Simopoulos AP: acidi grassi omega-3 nelle malattie di infiammazione e autoinnumma. J AM COLLUM 2002,21 No.6: 495-505.
5. Calder PC: grasso possibilità di immunomodulazione. Tendenze Immunol oggi 1996, 19, n. 6: 244-247.
6. Yaqoob P: Lipidi e la risposta immunitaria: dal meccanismo molecolare alle applicazioni cliniche. CURG AVAN NUTR Metab Assistenza 2003, 6: 133-150.
7. Tomobe Yi, Morizawa K, Tsuchida M, Hibino H, Nakano Y, Tanaka Y: Docosaeseaenoico acido dietetico sopprime l’infiammazione e gli immunoresponsabilità nella reazione di ipersensibilità contatti nei topi. Lipidi 2000, 35, n. 1: 61-69.
8. Anderson me: determinazione del glutatione e del glutatione disulfuolo in campioni biologici. Meth enzymol 1985, 113: 548-555.
9. Krawisz Ye, Sharon P, Stenson WF: dosaggio quantitativo per infiammazione intestinale acuta basata sull’attività di mileloperossidasi. Valutazione dell’infiammazione nei modelli di ratti e criceti. Gastroenterologia 1984, 87: 1344-1350.
10. Kelley DS, Warren JM, Simon VA, Bartolini G, Mackey Be, Erickson KL: effetti simili di C9, T11-CLA e T10, CL2CLA sulle funzioni di cellule immunitarie nei topi. Lipidi 2002, 37: 725-728.
11. Xaus J, Cardó M, Vellador AF, Solorer C. Lloveras J, Celada A: Interferon Gamma induce l’espressione di P21WAF1 e arresta il ciclo cellulare macrofagi che impedisce l’induzione dell’apoptosi. Immunità 1990, 2: 103-113.
12. PETERSON LD, THIES F, Sanderson P, Newsholme EA, PC calder: bassi livelli di acidi eicosapentaenoici e docosaesoici imitano l’effetto dell’olio di pesce sui linfociti del ratto. Vita SCI 1998, 62 (24): 2209-2017.
13. PETERSON LD, JEFFERY NM, THES F, Sanderson P, Newsholme EA Y Calder PC: Eicosapentaenoic e docosaeseenoic Acides Alter Rat milza Leukocyte acidi grassi Composizione e prostaglandin E2 Produzione ma hanno effetti diversi sulla funzione di linfociti e l’immunità mediata cellulare. Lipidi 1998, 33 (2): 171-180.
14. Thies F, Neve-Von-Caron G, Powell Jr, Yaqoob, P, Newsholme EA, PC calder: Integratore alimentare con acido gamma-linolenico o olio di pesce diminuisce la proliferazione dei linfociti in sano anziano. J Cutr 2001, 131 (7): 1918-1927.
15. Kew S, Benerjee T, Minhane Am, Finnegan Ye, Muggli R, Albers R, Williams cm, PC calder. Mancanza di effetti degli alimenti arricchiti da acidi grassi N-3 derivati da piante-ormarina sulle funzioni immunitarie umane. AM J CLIN CUL 2003, 77 (5): 1287-1295.
16. TREBBLE TM Y COLS.: PROSTAGLANDIN E2 Produzione e funzione cellulare T dopo l’integrazione di pesci pesci: risposta al cosupplementazione antiossidante. Am J Clinistruttore 2003, 78 (3): 376-382.
17. Endres S Y Cols.: L’effetto della supplementazione alimentare con gli acidi grassi N-3 sulla sintesi del fattore di Interleukin-1 e del fattore di necrosi tumorale da cellule mononucleari. N Engl J Med 1989, 320 (5): 265-271.
18. Calder PC, Yaqoob P, Thies F, Wallace Fa y Miles EA: acidi grassi e funzioni di linfociti. Brit J Cutr 2002, 87 supp per. 1, S31-S48.
19. Puertollano MA, De Pablo M, Álvarez de Cienfuegos G: Proprietà antiossidanti di N-acetil-cystcine Non migliorare la resistenza immunitaria dei topi nutriti lipidi dietetici a Listeria Monocytogenes infezione. Clinistro, 2003, 22 (3): 313-319.
20. Foitzik T, Eibl G, Schneider P, Wenger Fa, Giacobi Ca, Buhr HJ: IMEGA-3 L’integrazione di acidi grassi aumenta le citochine antinfiammatorie e attenua sequele di malattia sistemica in pancreatite sperimentale. Parentere Enteral Iutr, 2002, 26 (6): 351-356.
21. Wallace Fa, Miles Ea, Calder PC: Confronto degli effetti dell’olio di lino e delle diverse dosi di olio di pesce sulla funzione di cellule mononucleare in sani sottognioni umani. BR J CUST 2003, 89 (5): 679-681.
22. Hayashi Y Cols.: Effetti dell’emulsione di grasso omega-3 e omega-6 omega-6 sulla produzione di citochine e la sensibilità del tipo di navigazione in ritardo nei ratti bruciati che ricevono la nutrizione parenterale totale. J Parente ENTERAL GUST 1998, 22 (6): 363-367.
23. Raerlerstorff D, Pantze m, Bachmann H, Moser U: Proprietà antinfiammatorie degli acidi docosaeseenoici ed eicosupentaenoici nell’infiammazione dell’orecchio del mouse indotto da Phorbol-estere. Int Arch Allergy Immunol 1996, 111 (3): 284-290.
24. Jolly CA, Muthukumar A, Avula CP, Troyer D, Fernandes G: La durata della vita è progreditata nei topi Autoimmune-PRONE (NZBXNZW) resistenti al cibo (NZBXNZW) F (L) Fed a dieta arricchita con (N-3) acidi grassi. J Cutr 2001, 131 (10): 2753-60.
25. DEMOZ A, WILLUMSEN N Y BERGE RK: L’acido eicosapentaenoico alla dose ipotrigeridica migliora la difesa antiossidante epatico nei topi. Lipidi 1992, 27 (12): 968-971.