Originale
Alcuni aspetti della dinamica della popolazione degli ippopotami di Caranx ( Pesci: Carangidae) in Ash Bocas, Caribbean Colombiano
Alcuni aspetti della dinamica della popolazione del Crevalle Jack Catanx Ippopotami (Pesci: Carangidae) a Bocas de Ash, Caribbean Colombiano
1ministeriale di agricoltura e sviluppo rurale, Bogotá, Colombia.
2università della Magdalena, Programma di ingegneria della pesca, gruppo di biodiversità e ecologia applicata, Santa Marta, Maddalena, Colombia.
3Corporation Colombia International (JRC), Barranquilla, Atlántico, Colombia.
* Corrispondenza: [email protected]
ricevuto: settembre 2009; Accettato: luglio 2010.
Riepilogo
Obiettivo. Lo scopo di questo studio era di valutare gli aspetti biologici e la dinamica della popolazione degli Hollow (Caranx Hippos) nel settore Bocas Ash, dei Caraibi colombiani. Materiali e metodi. Tra giugno 2005 e giugno 2006, i dati di frequenza delle lunghezze commerciali sono stati raccolti per determinare il rapporto di pesatura della lunghezza (LH-W), la dimensione media della maturità sessuale (L50%), i parametri di crescita (K, L ∞ e T0) , tassi di mortalità (m, f e z) e sfruttamento (E). C’erano 1151 individui, di cui 264 sono stati utilizzati per l’analisi biologica. Risultati. Il rapporto LH-W differiva tra i sessi. Un L50% è stato stimato a 63,6 cm di lh. I parametri di crescita hanno presentato i seguenti valori: L∞ = 91 cm di LH, k = 0,38 anno-1 e a = 0,32 anni. Il valore dell’indice delle prestazioni ha avuto un intervallo compreso tra 3.33 e 3.73. Il tasso totale di mortalità (Z) era di 1,85 anni, la mortalità di pesca (f) di 0,72 anni e naturale (M) a 1,13 anni. Lo stato di sfruttamento (E) è stato di 0,30 anni. CONCLUSIONI. Secondo il tasso di sfruttamento, il giural non presenta sovrasplosentazione nelle bocche di cenere. Tuttavia, quando si confrontano la dimensione media della cattura con L50%, la specie viene acquisita con la principale attrezzatura da pesca prima del 50% della popolazione riesce a maturare. Linee guida per la progettazione di un sistema di pesca che garantisce la conservazione e l’uso sostenibile di questa risorsa sono proposti.
Parole chiave: pesca, riproduzione, ippopotami di carabro, caraibici, Colombia. (Fonte: obiettivi).
Astratto
Obiettivo. Per valutare gli aspetti biologici e le dinamiche della popolazione del Jack Crevalle (ippopotami di Caranx) da Bocas de Ash, dei Caraibi colombiani. Materiali e metodi. Tra giugno 2005 e il giugno dei dati di frequenza di lunghezza 2006 sono stati raccolti per determinare la relazione di peso della forcella, la dimensione media della scadenza (L50%), i parametri di crescita dell’equazione di Von BertanFaNFaNFY (K, L∞ e T0), la mortalità (m, f e z) e Tariffe di sfruttamento (e). Per un totale di 1151 individui sono stati campione, e 264 sono stati utilizzati per l’analisi biologica. Risultati La formazione di lunghezza della forcella ha mostrato differenze tra i generi. L50% è stato stabilito in 63,6 cm fl. I parametri di crescita I valori erano: L∞ = 91 cm fl, k = 0,38 anno-1 e a = 0,32 anni. I valori di indice di crescita comparativa (φ “) variavano da 3,33 a 3.73. Il tasso totale della mortalità (Z) è stato stimato come 1,85 anno-1, la mortalità di pesca (F) come 0,72 anni e la mortalità naturale (M) come 1,13 anno-1. Il tasso di sfruttamento (E) è stato determinato come 0,31 anni-1. CONCLUSIONI. Il tasso di sfruttamento ha indicato che Jack Crevalle da Bocass Bocas non è stato sovraesplosificato; Prima del 50% della popolazione raggiunge la sua maturità sessuale. Proponiamo di quadro per il sistema di gestione della pesca, assicurando la conservazione e l’uso adeguato di questa risorsa.
Parole chiave: pesca, riproduzione, ippopotami di carabro, caraibici, Colombia. (Fonte: obiettivi).
Introduzione
La famiglia Carangidae è uno dei gruppi di pesce della massima importanza economica nella pesca tropicale e subtropicale del mondo (1, 2). Dal landgoing dalla pesca artigianale nel dipartimento Atlantico, il Caranx Ippos (Linnae 1766) è una delle specie di maggiore domanda (3). Rappresenta il 47% nelle catture sbarcate annualmente e il 70% del reddito economico della popolazione pescatrice, essendo principalmente catturato con Translal e in alcuni casi con ganci e bowling (3). Nonostante questo, ci sono poche informazioni dettagliate sugli aspetti biologici e popolati che consentono di chiarire lo stato di sfruttamento in cui la sua popolazione si trova alla foce del fiume Maddalena.
Le stime dei parametri di crescita, della mortalità e dei tassi di funzionamento sono una parte importante nello studio della biologia dei pesci commerciali, in quanto consentono di determinare i livelli di sfruttamento e forniscono strumenti per l’attuazione delle politiche di amministrazione e di controllo per un adeguato sfruttamento (4-6). L’obiettivo di questo lavoro è stato quello di determinare il rapporto LH-W, la dimensione media della maturità sessuale, i parametri di crescita, i tassi di mortalità e di sfruttamento di C. ippopotami nelle Ash Bocas per conoscere lo stato di sfruttamento in questo settore e lo strumento scientifico Basi per proporre orientamenti orientati alla progettazione di sistemi di pesca che portano a uno sfruttamento sostenibile di questa importante risorsa.
Materiali e metodi
Area di studio. L’estuario del fiume Maddalena compreso nel Mar dei Caraibi, settore chiamato Ash Bocas (11 ° 06 ‘- 11 ° 08’ Nord; 74 ° 50 ‘- 74 ° 52’ OCCIA; Figura 1). Questo settore si trova a nord del Dipartimento Atlantico (3). Il clima è caratterizzato dalla presenza di due periodi climatici che influenzano l’attività dei pescatori (7); L’asciutto che inizia all’inizio di dicembre e finisce alla fine di aprile senza precipitazioni e una diminuzione del flusso del fiume Maddalena con una media di 4068 m3 S-1 (7, 8). Il periodo piovoso inizia a maggio e termina all’inizio di dicembre con una diminuzione dei venti e della nuvolosità elevata (7, 9) ed è quando vengono presentate le più grandi scariche d’acqua con una media di 10287 m3 s-1 (8).
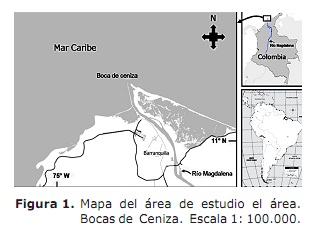
bocche di cenere è un estuario di soluzione salina, dove il suo funzionamento risponde all’interazione tra l’onda di marea e il contributo fluviale all’interno i contorni imposti dalla geometria dello stesso estuario (9). Il settore è influenzato da due correnti: i Caraibi dai Caraibi con un senso ovest e la corrente di Panama con questo (9). In questo settore, l’attività di pesca è di tipo fatta a mano e ha circa 250 pescatori che operano gli attrezzi da pesca, i ganci, le arti del bowling. Le catture sono rappresentate da specie marine, oltre a specie di sosta. Macrodon Ancyclodon, Caranx Crysos, C. ippopotami, Scomberomorus brasiliensis, ARIOPIS SP, SPP, SPP. e Arius Props (3).
Field e fase di laboratorio. Tra giugno 2005 e giugno 2006, i campionamenti sono stati tenuti per cinque giorni di ogni mese per registrare frequenze di lunghezze di catture commerciali atterrate nel quartiere di Las Flores, Barranquilla. Le catture sono state eseguite con ganci da calibro 7-10 e con ciotole e transazioni tra 8,89 e 20,32 cm di dimensioni della rete. Ogni individuo è stato sesso e pesante (senza eviscerazione) con un saldo di precisione di 0,1 g. Dal momento che non tutti i pesci sono arrivati con flussi completi pinne, è stata utilizzata la lunghezza della forcella. Le misurazioni sono state apportate al limite inferiore, utilizzando un Ichiometro con precisione di 0,1 cm (3). Per l’analisi biologica, le osservazioni macroscopiche dei gonadi sono state condotte per determinare il sesso e lo stato della maturazione gonadica, prendendo come riferimento la classificazione qualitativa del Vazzolelelelelelelere (10), che consiste di quattro stati: indifferenziati; immaturo (I); in maturazione (II); Maturo (III) e generato (IV).
Rapporto di lunghezza-peso (LH-W). Il rapporto di lunghezza-peso è stato determinato dal sesso attraverso la potenziale equazione w = ALHB, dove W è il peso totale, A e B sono costanti della potenziale regressione e LH è la lunghezza della forcella. Per determinare le differenze nel rapporto LH-W tra sessi, è stata eseguita un’analisi di covarianza (ANCOVA), dove la variabile dipendente era w, la variabile indipendente era lh e il fattore del sesso. Prima dell’analisi, sono state valutate le ipotesi di normalità (Kolmogorov-Smirnov), omogeneità delle varianze (Test Bartlett), rapporto lineare tra la variabile w e lh (analisi di correlazione) e omogeneità degli orecchini della relazione tra w e lh .
Mezza azione di maturità sessuale (L50%), composizione di cattura da taglie e dimensioni medi di acquisizione (TMC). Per valutare l’impatto della pesca sulla biologia di C. ippopotami, è stato calcolato L50%. Quegli esemplari i cui gonadi sono stati osservati negli stati III e IV sono stati considerati. Il calcolo era basato sul modello logistico che riguarda la proporzione di individui maturi con lunghezza (l; 5):
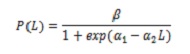
dove , β = 1; α1 e α2 sono i parametri della regressione.
Per stimare L50%, è stato utilizzato il rapporto -α1 / α2. Per regolare il modello, il metodo di regressione non lineare è stato utilizzato dai quadrati minimi, utilizzando l’algoritmo Iterativo Newton.
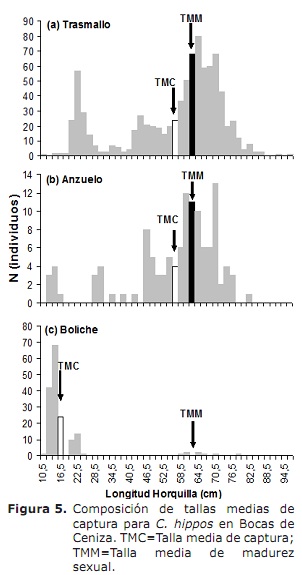
La composizione della cattura della dimensione è stata utilizzata per calcolare il TMC e i parametri di crescita.I dati sono stati raggruppati mensilmente nei compagni di classe ogni due centimetri. La relazione tra la frequenza relativa e il numero totale di individui è stato utilizzato per calcolare il TMC utilizzando la procedura proposta da Sparre e Venema (5); Successivamente, un TMC è stato calcolato per ciascuna arte di pesca osservata. Il TMC è stato confrontato con L50% per valutare l’impatto della pesca sulla biologia delle specie.
Parametri di crescita. I dati della lunghezza sono stati raggruppati mensilmente in compagni di classe di 1 cm. I parametri di crescita sono stati stimati da tre routine contenute nella versione computazionale FISAT versione 1.1 (4): l’analisi della frequenza elettronica (Elefan I), quella delle lunghezze di fogli di pastore (SLCA, 11) e il metodo di proiezione matrica (Projmat; 12) . La routine Elefan che collaboo con la crescita stagionalizzata dell’equazione di von Bertatanfy (Elefan I), essendo le dimensioni espresse in base all’età (5):
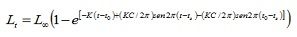
Dove, LT è la lunghezza per età T; La lunghezza asintotica; K è la costante di crescita, per il parametro di condizione iniziale, che determina il punto ipotetico nel tempo in cui il pesce ha una dimensione uguale a zero; C è il parametro di ampiezza dell’oscillazione della crescita; TS è parte dell’anno in cui il tasso di crescita è minimo.
La routine SLCA stima la crescita indipendente del numero e la posizione delle mode nel campione, in base all’impostazione della funzione Coseno:
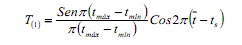
dove, Tmax e tmin sono età corrispondenti ai limiti superiore e inferiore di un determinato intervallo di classe; È l’età media e la TS è la frazione dell’anno in cui è stato preso il campione (6).
Il Projmat proietta un campione di distribuzione della frequenza delle lunghezze del tempo T + 1, utilizzando K e Valori L∞. Le frequenze proiettate sono state confrontate con quelle osservate dalla somma delle differenze dei quadrati, ripetendo il processo fino a ottenere le migliori stime (6).
tenendo conto del fatto che non è possibile ottenere una stima da parte delle frequenze di lunghezze (13,14), questo parametro è stato stimato dall’equazione empirica di Pauly (15):
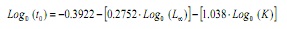
Dove, a è l’età ipotetica a cui l’animale inizia a crescere, la lunghezza asintotica e K è la costante della crescita
Per confrontare le stime di crescita tra i metodi, è stato utilizzato il tasso di prestazione della crescita φ “(16). Inoltre, questi valori sono stati confrontati con le stime di altre popolazioni di C. Ippopotami documentati in letteratura. Questo indice è stato stimato dall’espressione:
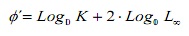
dove, k e l∞ sono i parametri di crescita.
Dato che il valore stimato di L∞ in questo studio era basato su LH e quelli documentati in letteratura in lunghezza totale (LT), a fini di fare confronti tra i valori, una semplice regressione lineare è stata effettuata tra LT (dipendente) e lh (indipendente) per rendere la conversione di lh a lt. Per questo il seguente modello lineare è stato ottenuto con i dati di questo studio:
lt = – 0.4387 + (1,2335 lh)
Dalle frequenze LH di catture commerciali, la mortalità totale (Z), naturale (M) e pesca (F) della popolazione di C. Ippos è stato proceduto a stimare. Z è stata stimata dalla curva di cattura linearizzata convertita in formati (17):
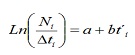
dove, né il numero di individui catturati appartenente alla classe di lunghezza I; T’i è l’età relativa della classe I; ΔTI è il tempo medio richiesto dall’individuo per la sua crescita attraverso la classe I.
m è stato stimato utilizzando l’equazione empirica di Paully (17):
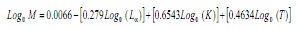
Dove, L∞ e K sono i parametri del von Bertalanffy e T Equation è la temperatura media annuale di acqua costiera di superficie nell’area di studio, che è stata considerata a 28.2 ° C. / P>
Il tasso di mortalità di pesca e il tasso operativo sono stati stimati da:
Per eseguire le stime dei parametri, è stata assunta una crescita stagionale di C. Ippos, dato che nel corso dell’anno, episodi ambientali nell’area di studio che potrebbero influenzare la crescita del pesce. Tale è il caso delle condizioni stagionali del caudale del fiume Maddalena, che può influenzare le condizioni fisiche, chimiche e biologiche a sua bocca (8). A questo è aggiunto, l’influenza dei venti di alisio nel settore della bocca di Ash, poiché quando sono in corso e secondo la loro intensità, cambiano anche le condizioni oceanografiche del settore (18). Allo stesso modo, la conoscenza biologica delle specie indica che C.Ippopotami è una specie migratoria (1), da cui la stagionalità può anche essere dedotta nella crescita. Sulla base di quanto sopra, i valori trovati da Elefan sono stati selezionati per stimare i tassi di mortalità di C. ippopotami in questo studio.
Con lo scopo di quantificare l’effetto dei campioni mensili nei parametri di crescita e Ottenere i valori di accuratezza di ciò, è stato applicato la tecnica di remphing di jackknife (6). Ciò consiste nella stima delle dimensioni mensile n delle dimensioni della frequenza delle dimensioni e sequenzialmente vengono omesse per ottenere combinazioni di ciascuno dei parametri. Per specificare questa stima, è stato utilizzato l’errore percentuale (EP) e il coefficiente di variazione (CV), utilizzando le seguenti formule:
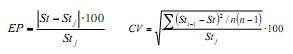
Dove, ST è il parametro di crescita stimato utilizzando il full banca dati annuale e STJ è lo stimatore medio derivato dalle successive stime del jackknife; STI-1 è il parametro stimato quando si omettono il mese I, e N è la dimensione del campione annuale (n = 12).
Risultati
Durante il periodo di studio sono stati raccolti individui, di cui 868 sono stati catturati con Translal, 120 con un gancio e 167 con bowling; Presentando una gamma di dimensioni tra forcella lunga 10,5 e 96,5 cm. Per l’analisi degli aspetti biologici studiati, sono stati conteggiati 264 individui, essendo 180 maschi e 84 femmine, con una serie di dimensioni tra 25 e 96,5 cm di LH e un peso totale tra 281,5 e 10500 g.
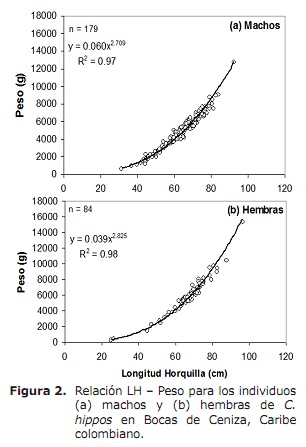
Rapporto di lunghezza-peso (LH-W). L’analisi ha rivelato che tra il sesso del rapporto LH-W differiva in modo significativo, osservando che le femmine pesano più dei maschi alla stessa lunghezza (ANCOVA: F1, 260 = 21.5, P < 0.05 ). Pertanto, il rapporto LH-W per maschi era w = 0,060LH2.709 (P < 0,05) e per le femmine w = 0,039LH2.825 (P < 0.05) (figura 2). I valori di B derivanti da regressioni per entrambi i sessi erano significativamente inferiori a 3, indicando una crescita minore (Test T, P > 0,05; 15).
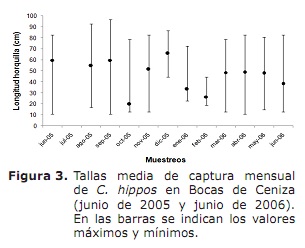
composizione della dimensione, dimensione media della maturità sessuale (L50%) e dimensioni medi di acquisizione (TMC). In generale, le dimensioni minime di acquisizione sono state registrate in ottobre, novembre 2005 e marzo, aprile e giugno 2006; Mentre le massime erano frequenti durante tutto il periodo di studio, tranne per ottobre, gennaio e febbraio (figura 3).
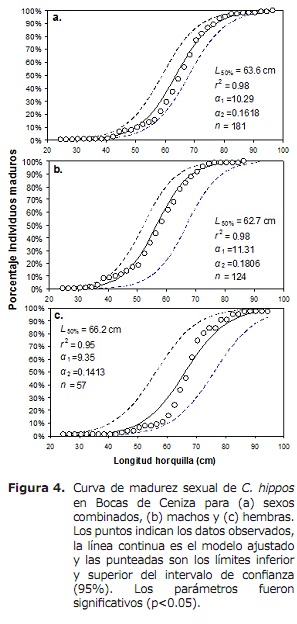
l50% per i sessi combinati era 63,6 cm LH (95% CI: 59,0-68,4 cm). Per il caso di maschi e femmine erano 62,6 cm (95% CI: 58.3-70 cm) e 66,2 cm (95% CI: 56,1-76,2 cm), rispettivamente (Figura 4). Il TMC globale è stato stimato a 50,3 cm di LH. Per il caso di transazioni, i ganci e il bowling sono stati stimati a 56,0, 56,2 e 17,8 cm, rispettivamente (figura 5A, 5b e 5c), sotto L50%.
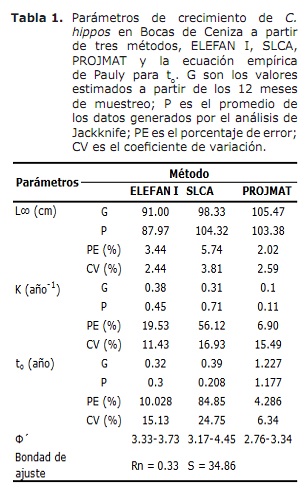
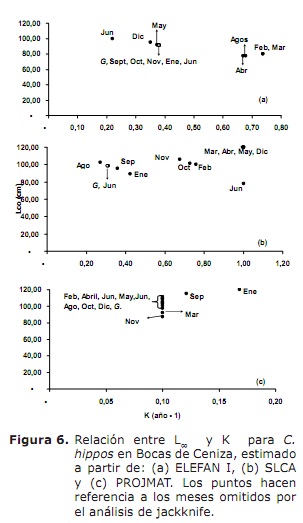
crescita. I risultati di L∞ e K forniti dai metodi Elefan I, SLCA e Projmat sono presentati nella Tabella 1. Si è osservata che questi tre metodi hanno fornito valori diversi per ciascuno dei parametri di crescita. L’analisi di Jackknife ha mostrato che esiste un effetto dei dati sui diversi parametri di crescita, in particolare per SLCA e Projmat. Per il caso di Elefan I, L∞ variava da 77,4 a 100 cm, mentre i valori K erano tra 0,20 e 0,75 anni-1 (figura 6). Con SLCA, i valori più alti di L∞ e K sono stati ottenuti con valori tra 78,25 e 120 cm e rispettivamente compreso tra 0,3 e 1,0 anni. Secondo i valori gettati da Projmat, è stata osservata meno dispersione per K fluttuante tra 0,10 e 0,17 anni-1. Il contrario si è verificato per L∞, dove i valori variavano tra 87 e 120 cm, essendo quest’ultimo superiore a quello osservato nei precedenti stimatori di crescita. I valori K stimati hanno mostrato differenze tra i metodi, con i più alti quelli calcolati da Elefan I e SLCA (Tabella 1). Il valore ottenuto per l’età ipotetica a cui l’animale (A) inizia a crescere dall’equazione empirica di Paully (20) è stata di 0,32 anni, con un coefficiente di variazione del 13,92% e una percentuale di errore (PE) del 10.02. L’analisi dell’indice di performance ǿ ha mostrato valori simili tra Elefan I (3.33-3.73) e SLCA (3.17-4.45), che non si è verificato per Projmat che ha mostrato valori inferiori rispetto ai precedenti (2,76-3.34). Anche così, la variazione inferiore è stata presentata per Elefan I, seguita da Projmat e Slca, quest’ultimo è il rango più alto presentato. Tenendo conto del coefficiente di variazione (CV) di ciascuno dei parametri, è stato rilevato che le stime Elefan I delle analisi di Jackknife tendevano a fornire valori inferiori per L∞, a differenza di SLCA e Projmat che hanno raggiunto valori più elevati (tabella 1) .Il risultato del PE ottenuto per K con Projmat era inferiore a quanto trovato per Elefan I e Slca. Di conseguenza, il metodo che ha gettato le stime più credibili dei parametri di crescita è stata Projmat, seguita da Elefan I e infine SLCA.
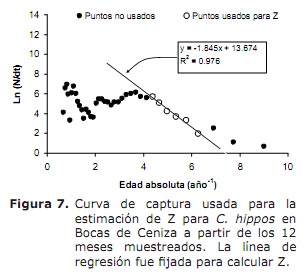
mortalità e stato di sfruttamento. Basato sui seguenti parametri di crescita: k = 0,38 anno-1; L∞ = 91 cm lh; a = 0,32 anni; Oscillazione stagionale nel modello di crescita (c) di 0,5 e un tasso di crescita minimo (WP) di 0,3, è stato stimato un tasso totale di mortalità (Z) di 1,76 anni (figura 7), oscillante tra 1,38 e 2,22 anni -1 ( dall’analisi di Jackknife).
La mortalità naturale (M) era 1,12 anno-1, variabile tra l’1,10 e 1,14 anni. Il tasso di mortalità di pesca (F) è stato stimato a 0,63 anni, presentando una serie di variazioni comprese tra 0,2 e 1,06 anni. I valori del rapporto F e Z per stimare il tasso operativo (E), hanno presentato un valore massimo di 0,5 anni, in media di 0,30 anni.
Discussione
Rapporto di lunghezza-peso (LH-W). Il concetto di crescita comporta cambiamenti relativi delle dimensioni e del peso degli individui (19), e la loro relazione ha diversi usi nella ricerca di pesca. Ad esempio, è utile nei modelli di analisi delle scorte di pesca per stimare la biomassa di una popolazione; per conoscere le condizioni di un pesce; Per confrontare la storia della vita di determinate specie tra regioni; e determinare il tipo di crescita che il pesce di solito presente (20). C. Ippos, presenta un peso senza peso del tipo di tipo alometrico (< 3; 15), indicando che presenta tassi di crescita differenziali per diverse parti del corpo, dove la lunghezza aumenta a A un tasso più alto del peso.
I risultati indicano che questa condizione era più evidente per i maschi che per le femmine, dato che quest’ultimo, nonostante la presentazione di una crescita elementare minore, approssimativa avere una crescita isometrica. Questo è stato concordante con il risultato dell’Ancova, che ha indicato che lo stesso lh ci sono differenze nel peso sessuale. Le femmine di questa specie presentano maggiore peso e altezza del corpo rispetto ai maschi. D’altra parte, è importante evidenziare che, poiché i campioni sono stati raccolti per un anno, il rapporto LH-W in questo studio non rappresenta un particolare periodo dell’anno. In questo senso, i valori B potrebbero essere considerati un valore medio annuo per la specie (20). I risultati di questa ricerca erano simili a quelli documentati per le specie in diverse aree geografiche. Ad esempio, i valori B-basati su B sono stati documentati da 2,73 nel sud della Florida (21), dal 2,85 in India (1) e 2,91 nel Golfo di Salamanca, in Colombia (22).
Crescita, mortalità e stato di sfruttamento. Sulla base dei parametri di crescita ottenuti da Elefan I, la lunghezza massima stimata per C. Ippos in Ash Bocas era 91 cm LH corrispondente a 111,81 cm di lt. Se quest’ultimo viene convertito in età corrispondente a 14 anni, ottenuto con un tasso di crescita K di 0,38 anni-1, suggerendo che C. Ippos è una specie longeva (17). D’altra parte, tenendo conto della curva della crescita, questa specie è cresciuta rapidamente fino all’età di 5 anni. Questi dati coincidono per altri archiviazioni, come quella di Trachinoto Falcatus, che raggiunge una dimensione massima di 90 cm e ha anche una crescita massima di 5 anni, da cui avvia la fase stazionaria della sua crescita (2).
D’altra parte, i valori di L∞ stimati per C. Ippos in questo studio erano superiori a quelli documentati in letteratura; Mentre quelli dell’indice di performance ǿ rivisto nella letteratura hanno mostrato valori molto vicini alla stima in questo studio (tabella 2). Assumere questo indice come criterio di confronto tra i valori dei parametri di crescita ottenuti e tenendo conto che C. Ippos è una specie migratoria (23), si può presumere che la crescita di questa specie sia concordanza in quanto descritto in diverse aree della sua distribuzione globale (tabella 2).
Il modello di crescita annuale di C. ippopotami ha mostrato oscillazioni (c = 0.5) Con un tasso di crescita minimo nel mese di settembre (WP = 0.3). Questo modello può essere influenzato da uno degli aspetti più caratteristici della crescita del pesce: la variabilità osservata tra individui di una popolazione e tra le popolazioni di una specie, che è direttamente correlata all’ambiente (24). Ad esempio, la variabilità delle condizioni fisico-chimiche dell’acqua (24) e della qualità e della quantità di cibo (19) condiziona la migrazione di una specie, che comprende gli spostamenti della stazione tra le zone di allevamento, alimentazione e deposizione delle uova (15, 25) .
Sulla base di quanto sopra e tenendo conto delle frequenze delle lunghezze osservate durante il periodo di studio, che indicano anche la possibile presenza di stagionalità nella crescita (13), può essere dedotta che la crescita di C. ippopotami è stagionale. Per quanto riguarda il tasso di crescita minimo, questo evento potrebbe essere dato dalla fase riproduttiva della specie, poiché entro settembre è stato osservato un aumento del numero di individui maturi (nelle fasi III).
D’altra parte, la relazione tra il tasso di mortalità di pesca (f) e la mortalità totale (Z) di 0,3, indica che la popolazione nell’area delle bocche di cenere non è in esecuzione il rischio di essere sovraesplorato , sotto la premessa che una popolazione raggiunge le sue prestazioni ottimali quando la frazione di morte causata da F è superiore a 0,5 (6,13). Tuttavia, è importante sottolineare che quando L50% è stato paragonato a TMC, C. Ippopotami hanno presentato una maggiore pressione sulla popolazione adulta, catturata frequentemente con trasmissione e gancio. La situazione peggiora con l’uso di Boliche, che non solo colpisce gli adulti, ma anche i giovani, che non consentono il rinnovamento naturale di questa risorsa di pesca.
Proposta per la progettazione di un sistema di gestione della pesca . A partire dalla base che la pesca eccessiva può avvenire in tre modi diversi: (i) in relazione alla crescita; (ii) rispetto al reclutamento; e (iii) sovrastaccare che colpisce l’ecosistema (15). In questo senso, la pesca di C. ippopotami nel settore della bocca di Ash può essere classificata all’interno del primo oggetto, che avviene quando gli individui vengono catturati prima che raggiungano le dimensioni necessarie per garantire il rinnovamento biologico della popolazione. Quanto sopra è diventato un problema comune per la maggior parte delle specie commerciali ed è stato documentato per molte specie costiere nei Caraibi colombiani.
Tutto quanto sopra, è necessario che gli sforzi invesentano per progettare la gestione delle strategie di progettazione per C. Ippopotami nella pesca di ASHO Bocks, in modo tale da essere coerenti con l’approccio precauzionale al pescatore responsabile di questa risorsa (27). Caddy (27) suggerisce che le strategie debbano essere applicate all’interno di un quadro di ridondanza di gestione in cui tale quadro potrebbe consistere in una miscela di misure che coinvolgono arti, criteri di secking temporanei e aumento delle dimensioni minime di acquisizione. In questo senso, tenendo conto del L50% come punto di riferimento limite (28), qui si consiglia di fissare la dimensione minima di acquisizione di questa risorsa a forchetta lunga 64 cm. Allo stesso modo, in tutti gli ingranaggi da pesca utilizzati, si propone di aumentare le dimensioni della rete superiore a 8,89 cm e utilizzare i calibri del gancio finalizzati a catturare pesci più grandi. Nelle condizioni attuali, queste attrezzature da pesca interessano direttamente la frazione di giovani individui che entrano nella prima fase della maturità sessuale. Con le misure di cui sopra, integrate con altre variabili come la cattura, lo sforzo di pesca e la cattura per unità di sforzo, così come fattori sociali ed economici, si prevede che C. Ippopotassino sia catturato in una maggiore proporzione di individui sopra di L50% garantire la sostenibilità di questa risorsa nel tempo. In questo senso, è importante tenere conto dello stato multispecifico e multiflot della pesca e pertanto, è raccomandato un processo di accordo tra i pescatori, i commercianti e le entità pertinenti per raggiungere accordi volti a uno sfruttamento responsabile della risorsa.
In conclusione, la Jorel non presenta sovrasplosentazione in bocche di cenere. Tuttavia, quando si confrontano la dimensione media della cattura con L50%, la specie viene acquisita con la principale attrezzatura da pesca prima del 50% della popolazione riesce a maturare. Linee guida per la progettazione di un sistema di pesca che garantisce la conservazione e l’uso sostenibile di questa risorsa sono proposti.
ACCONDICI
Alla comunità dei pescatori, membri di Asopscar e Coopez. Alla Brothers Villa e La Pescadería El Nuevo Tambó per la sua collaborazione nel coordinamento delle attività e della fornitura di informazioni sul campo. Agli insegnanti G. Lozano, L.C. Gutiérrez, O. Galvis, J. Bianco ed E. Viloria.
Riferimenti
1.Ruben, S, HM Kasim, S Sivakami, PN Radhakrihnan, Kn Kurup, M Sivila, Noble, KVS Nair, Raje SG. Pesca, biologia e valutazione delle scorte delle risorse Carangid dai mari indiani. Indian J Fish 1992; 39 (3,4): 195-234.
2. Crabree, R, PB Hood, Snodgrass D. Età, Grouth e riproduzione del permesso (Trachinotus Falcatus) nelle acque della Florida. Pesce Bull 2002; 100: 26-34.
3.River R, Solano, G.Diagnosi dell’attività di pesca artigianale nella zona costiera del dipartimento Atlantico, dei Caraibi colombiani. Barranquilla, Colombia: Incoder; 2005.
4.Gayalino, FC, P sparso, Paully D. La Guida dell’utente FISAT. FAO – Strumenti ICLARM Stock Strumenti. Roma: FAO; 1993.
5.Sparre, P., Venema S. Introduzione alla valutazione delle risorse di pesca tropicale. Roma: FAO; 1995. Documento tecnico della pesca: 306/1.
6.Rueda, M, M, Santos-Martínez A. Dinamica della popolazione del Mujarra a strisce Eugerres Plumieri dal grande Ciénaga de Santa Marta, in Colombia. Pesce res 1999; 42: 155-166.
7.banco, Ja, EA Viloria, Narváez BJC. ENSO e cambiamenti di salinità interinnuali nel grande sistema laguna del Coastal Ciénaga de Santa Marta, Caraibi colombiani. Shelf SCI di ESTUAR Coast SCI 2006; 66: 157-167.
8. RESTEPO, JD, KJERFVE B. MAGDALENA River: variabilità interinnica (1975-1995) e revisionata scarico dell’acqua e stime del carico dei sedimenti. J Hydrol 2000; 235 (1-2): 37-149.
9.ACEVEDO A, GUTIÉRREZ ML. Alcuni aspetti riproduttivi del rizooonodon poroso Toyo (1831) (Elasmobranchii: Carccharinidae). Nel settore della bocca di Ash, dei Caraibi colombiani. . Barranquilla, Colombia: Atlantic University, Facoltà di Scienze di base; 1999.
10.VazzoleleLer, EAM. Manuale del metodo per Popolare Pexes Populare Studisti biologici: riproduzione e crescimento. Brasília: CNPQ; 1982.
11.sheperd, JG. Un metodo debolmente parametrico per l’analisi dei dati della composizione della lunghezza. In: Paully, D, editori GP di Morgan. Metodi a base di lunghezza nella ricerca della pesca. Manila, Iclarm Conference Procedings 13. 1987.
12.lielie, pH. L’uso di matrici in certa matematica della popolazione. BIOMETRIKA 1945; 35: 213-245.
13.Gulland, Ja, Rosemberg AA. Esame dai metodi basati sulle dimensioni per valutare le popolazioni di pesci. Roma: FAO; 1992.
14.toRdecilla-Petro G, S Sánchez-Band, Olaya-Nieto Ch. Crescita di Moncholo e mortalità (Hoplias Malabaricus) al Lorica Lorica, in Colombia. Rev mvz córdoba 2005; 10 (2): 623-632.
15.Auly, D. Alcuni semplici metodi per la valutazione delle risorse tropicali della pesca. Roma: FAO; 1983.
16.PAY, D, MUNRO JL. Undici di più sul confronto della crescita dei pesci e degli invertebrati. Fishb 1984; 1:21.
17.Auly, D. Dinamica della popolazione del pesce nelle acque tropicali; Un manuale per l’uso con calcolatori programmabili. Iclarm Stud Rev 1984; 8: 235.
18.blanco, ja. Variazioni ambientali stagionali nelle acque costiere e la sua importanza per la pesca nella Santa Marta, nella regione dei Caraibi colombiani. . Santa Marta-Bogotá. Università nazionale della Colombia, Facoltà di Scienze di base; 1988.
19.abaunza L, Karlou-Riga C, Murta A, Eltink W, García Santamaría Mt, Zimmermman c, martello c, et al. Crescita e riproduzione dello sgombro del cavallo, Trachurus Trachurus (Carangidae). REV PESCIO BIOOL PESCE 2003; 13: 27-61.
20.Froese, R. Cube Law, Condition Factor e Bisce-Lunghezza Relazione: Storia, Meta-analisi e raccomandazioni. J appl iththyol 2006; 22: 241-253.
21.bohsack, ja, harper de. Le relazioni di peso di lunghezza della barriera corallina marina selezionata pesca dagli Stati Uniti sud-orientale e dai Caraibi. Memorandum tecnico NOAA; 1988. NMFS-SEFC-215.
22.Duarte, Lo, CB García, I Moreno, G Melo, P Navajas, N Sandoval, et al. Atlante demografico del Golfo di Salamanca, Caraibi colombiani: dinamica della popolazione, distribuzione, cibo e riproduzione. . Versione 1. Santa Marta: Invemar-UNAL-Colciencias; 1999.
23.HoneyBrink, R. Una revisione della biologia della famiglia Carangidae whith Empassis sulle specie trovate nelle acque hawaiane. Honolulu: Divisione delle risorse acuatiche; 2000.
24.macer, ct. Alcuni aspetti della biologia dello sgombro del cavallo (Trachurus Trachurus (l)) nelle acque in tutta la Gran Bretagna. J Fish Biol 1977; 10: 51-62.
25.harden, fr. Migrazione del pesce. Londra: Arnold; 1968.
26.Narváez B, JC, FRera, Bianco J. Effetti dell’ingranaggio della pesca sulle dimensioni del pesce in una pesca artigianale caraibica colombiana. BOL INVESTION MAR COSTA 2008; 37 (2): 163-187.
27.Caddy, JF. Gestione della pesca nel ventunesimo secolo: si applicano nuovi paradigmi?. Rev pesch Biol Pesce 1999; 9: 1-43.
28.Caddy, JF. Limitare punti di riferimento, semafori e approcci olistici alla gestione della pesca con ingresso minimo di valutazione delle scorte. Pesce res 2002; 56 (2): 133-137.
29.Torres, FJR, MBTM Norizam, LR Garcés, Silvestre GT. Compilazione dei parametri di popolazione di specie di pesci comunemente catturati in reciproci nel sud e nel sud-est asiatico. Manila: centro di pesce mondiale; 2001.
30.snelson, ff. Studi biologici di Crevalle Jack (ippopotami di Caranx) in Florida. St. Petsburg: Florida Marine Resesical Institute; 1992.