Original
Algúns aspectos da dinámica da poboación dos hipopótamos de Caranx ( Peixes: Carangidae) en Bocas de Ash, Caribe Colombiano
Algúns aspectos da dinámica da poboación do Crevalle Jack Caranx Hippos (Piscis: Carangidae) en Bocas de Ash, Caribe Colombiano
2Universidade da Magdalena, Programa de Enxeñaría Pesca, Grupo de Biodiversidade e Ecoloxía Aplicada, Santa Marta, Magdalena, Colombia.
3Corporation Colombia International (JRC), Barranquilla, Atlántico, Colombia.
* Correspondencia: [email protected]
Recibida: setembro de 2009; Aceptado: xullo de 2010.
Resumo
Obxectivo. O obxectivo deste estudo foi avaliar aspectos biolóxicos e a dinámica da poboación dos oco (Caranx Hippos) no sector de Bocas de Ash, o Caribe Colombiano. Materiais e métodos. Entre xuño de 2005 e xuño de 2006, recolléronse datos de frecuencia de lonxitudes comerciais para determinar a ratio de peso da lonxitude (LH-W), o tamaño medio da madurez sexual (L50%), os parámetros de crecemento (k, l ∞ e t0) , Taxas de mortalidade (M, F e Z) e Explotación (E). Había 1151 individuos, dos cales 264 foron utilizados para a análise biolóxica. Resultados. A relación LH-W difería entre os sexos. Un L50% estimouse en 63,6 cm de LH. Os parámetros de crecemento presentaron os seguintes valores: L∞ = 91 cm de LH, k = 0,38 ano-1 e = 0,32 ano. O valor do índice de rendemento tivo un intervalo entre 3.33 e 3.73. A taxa de mortalidade total (Z) foi de 1,85 anos, 1, mortalidade de pesca (F) de 0,72 anos-1 e natural (m) a 1.13 ano-1. O estado de explotación (E) foi de 0,30 anos-1. Conclusións Segundo a taxa de explotación, o xurelito non presenta a sobreexplotación en boca de cinza. Non obstante, ao comparar o tamaño medio de captura co L50%, a especie está sendo capturada cos principais artes de pesca antes do 50% da poboación consegue madura. Pautas para o deseño dun sistema de pesca que garante a conservación e uso sostible deste recurso son propostas.
Palabras clave: Pesca, reprodución, Caranx Hippos, Caribe, Colombia. (Fonte: Obxectivos).
Resumo
Obxectivo. Para avaliar os aspectos biolóxicos ea dinámica da poboación do Jack Crevalle (Caranx Hippos) de Bocas de Ash, o Caribe Colombiano. Materiais e métodos. Entre xuño de 2005 e os datos de frecuencia de xuño de 2006 foron recollidos para determinar a relación de lonxitude de tenedor, o tamaño medio de vencemento (L50%), os parámetros de crecemento da ecuación de von Bertalanffy (K, L∞ e T0) e Mortalidade (M, F e Z) e Taxas de explotación (e). Ao total de 1151 individuos foron a mostra, e 264 foron utilizados para a análise biolóxica. Resultados A relación de lonxitude de fork-peso mostrou diferenzas entre os xéneros. O L50% estaba estado en 63,6 cm. Os valores dos parámetros de crecemento foron: L∞ = 91 cm fl, k = 0.38 ano-1 e a = 0,32 ano. O índice de crecemento comparativo (φ ‘) valores variaron de 3,33 a 3,73. A taxa de mortalidade total (Z) foi estimada como 1,85 anos-1, mortalidade de pesca (f) como 0,72 anos-1 e mortalidade natural (m) como 1,13 ano-1. A taxa de explotación (E) foi determinada como 0,31 ano-1. Conclusións A taxa de explotación indicou que Crevalle Jack de Ash Bocas non estaba sobreexploited; Antes do 50% da poboación alcanza a súa madurez sexual. Propoñemos ao marco do sistema de xestión da pesca, asegurando a conservación e uso adecuado deste recurso.
Palabras clave: Pesca, reprodución, Caranx Hippos, Caribe, Colombia. (Fonte: Obxectivos).
Introdución
A familia de Carangidae é un dos grupos de peixes de maior importancia económica nas pesquerías tropicais e subtropicais do mundo (1, 2). Desde a terra da pesca artesanal no departamento atlántico, os hipopótamos de Caranx (Linnae 1766) é unha das especies de maior demanda (3). Representa o 47% en capturas desembarcadas anualmente eo 70% dos ingresos económicos da poboación pesquera, sendo capturado principalmente con transmisas e nalgúns casos con ganchos e bolos (3). Malia isto, hai pouca información detallada sobre os aspectos biolóxicos e de poboación que permiten elucidar o estado de explotación na que se atopa a súa poboación na desembocadura do río Magdalena.
As estimacións dos parámetros de crecemento, a mortalidade e as taxas de operación son unha parte importante no estudo da bioloxía de peixe comercial, xa que permiten determinar os niveis de explotación e proporcionan ferramentas para a implementación de políticas de administración e control para a explotación adecuada (4-6). O obxectivo deste traballo foi determinar a relación LH-W, o tamaño medio da madurez sexual, os parámetros de crecemento, a mortalidade e as taxas de explotación de C. hipopótamos en Ash Bocas para coñecer o estado de explotación neste sector e instrumento o científico Bases para propoñer directrices orientadas ao deseño de sistemas de pesca que leven a unha explotación sostible deste importante recurso.
Materiais e métodos
Área de estudo. O estuario do río Magdalena entendido no mar Caribe, o sector chamado Bocas de Ash (11 ° 06 ‘- 11 ° 08’ Norte; 74 ° 50 ‘- 74 ° 52’ West; Figura 1). Este sector está situado ao norte do Departamento Atlántico (3). O clima caracterízase pola presenza de dous períodos climáticos que afectan a actividade dos pescadores (7); O seco que comeza a principios de decembro e remata a finais de abril sen choiva e unha diminución do fluxo do río Magdalena cunha media de 4068 M3 S-1 (7, 8). O período de choiva comeza en maio e remata a principios de decembro cunha diminución dos ventos e a alta nubidade (7, 9) e é cando se presentan as maiores descargas de auga cunha media de 10287 M3 S-1 (8).
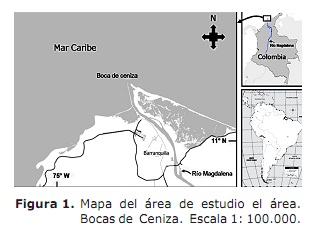
boca de cinzas é unha ría de salina, onde a súa operación responde á interacción entre a onda de marea ea contribución fluvial dentro Os contornos impostos pola xeometría do mesmo estuario (9). O sector está influenciado por dúas correntes: o Caribe do Caribe con sentido oeste e a corrente de Panamá con este (9). Nesta área, a actividade pesqueira é un tipo artesanal e ten preto de 250 pescadores que operan os artes de pesca, ganchos, lonxitude e artes de bolos. As capturas están representadas por especies mariñas, así como a supresión de especies. Macrodon Ancyclodon, Caranx Crysos, C. Hippos, Scomberomorus Braziliensis, Ariopis SP, SPP, SPP. e Arius Props (3).
Fase de campo e laboratorio. Entre xuño de 2005 e xuño de 2006, as mostras celebráronse durante cinco días de cada mes para gravar frecuencias de lonxitudes de capturas comerciais desembarcadas no barrio de Las Flores, Barranquilla. As capturas realizáronse con 7-10 ganchos de gauge e con cuncas e transmases entre 8,89 e 20,32 cm de tamaño de malla. Cada individuo foi sexual e pesado (sen evisceración) cun saldo de precisión de 0,1 g. Xa que non todos os peixes chegaron con aletas de fluxos cheos, utilizouse a lonxitude do garfo. As medidas foron feitas ao límite inferior, usando un ichtiometer con precisión de 0,1 cm (3). Para a análise biolóxica, realizáronse observacións macroscópicas das gónadas para determinar o sexo e estado da maduración gonadal, tomando como referencia a clasificación cualitativa de Vazzoleler (10), que consta de catro estados: indiferenciados; inmaduro (I); en maduración (II); Maduro (III) e xerado (IV).
Relación de peso de lonxitude (LH-W). A relación de peso de lonxitude foi determinada polo sexo a través da ecuación potencial W = Alhb, onde W é o peso total, A e B son constantes da regresión potencial e LH é a lonxitude do fork. Para determinar as diferenzas na relación LH-W entre sexos, realizouse unha análise de covarianza (ANCOVA), onde a variable dependente foi W, a variable independente foi LH e o factor de sexo. Antes da análise, avaliáronse as suposicións de normalidade (Kolmogorov-Smirnov), a homoxeneidade das varianzas (Bartlett Test), a relación lineal entre a variable W e LH (análise de correlación) e homoxeneidade dos pendentes da relación entre w e lh ..
Half stock de madurez sexual (L50%), composición de captura por tamaños e tamaño de captura media (TMC). Avaliar o impacto da pesca sobre a bioloxía de C. HIPPOS, calculouse o L50%. Eses especímenes cuxas gonadas foron observadas nos Estados III e IV foron considerados. O cálculo baseouse no modelo de loxística que relaciona a proporción de individuos maduros con lonxitude (l; 5):
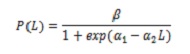
onde , β = 1; α1 e α2 son os parámetros da regresión.
Para estimar o L50%, usouse a relación -α1 / α2. Para axustar o modelo, o método de regresión non lineal foi utilizado por prazas mínimas, utilizando o algoritmo iterativo de Newton.
A composición da captura de tamaño foi utilizada para calcular o TMC e os parámetros de crecemento.Os datos foron agrupados mensualmente nos compañeiros de clase cada dous centímetros. A relación entre a frecuencia relativa eo número total de individuos utilizáronse para calcular a TMC usando o procedemento proposto por Sparre e Venema (5); Posteriormente, un TMC foi calculado para cada arte de pesca observada. O TMC foi comparado con L50% para avaliar o impacto da pesca sobre a bioloxía das especies.
parámetros de crecemento. Os datos de lonxitude foron agrupados mensualmente nos compañeiros de clase de 1 cm. Os parámetros de crecemento foron estimados por tres rutinas contidas no paquete computacional de Fisat versión 1.1 (4): a análise de frecuencia electrónica (Elefan I), a partir de lonxitudes de folla de pastor (SLCA; 11) eo método de proxección matriculado (projmat; 12) .. A rutina Elefan I traballa coa ecuación de crecemento estacionalizada de Von Bertalanffy (Elefan I), sendo o tamaño expresado de acordo coa idade (5):
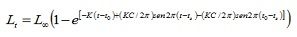
Onde, Lt é a lonxitude de idade T; L∞ a lonxitude asintótica; K é a constante de crecemento, é o parámetro de condición inicial, que determina o punto hipotético no tempo en que o peixe ten un tamaño igual a cero; C é o parámetro de amplitude do crecemento oscilación; TS forma parte do ano en que a taxa de crecemento é mínima.
A rutina SLCA estima o crecemento independente do número e posición das modas na mostra, en función da configuración de funcións do coseno:
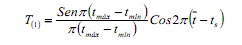
Onde, TMAX e TMIN son idades correspondentes aos límites superior e inferior dun intervalo de clase dado; É a idade media e TS é a fracción do ano en que se tomou a mostra (6).
O projmat proxecta unha mostra de distribución de frecuencia de tempo de lonxitude T + 1, usando o k e Valores de L∞. As frecuencias proxectadas foron comparadas coas observadas pola suma das diferenzas dos cadrados, repetindo o proceso ata obter as mellores estimacións (6).
tendo en conta que non é posible obter unha estimación de A partir das frecuencias de lonxitudes (13,14), este parámetro foi estimado a partir da ecuación empírica de Pauly (15):
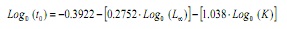
Onde, a é a idade hipotética á que o animal comeza a crecer, L∞ A lonxitude asintótica e K é a constante de crecemento
Para comparar estimacións de crecemento entre métodos, utilizouse a taxa de rendemento do crecemento φ ‘(16). Adicionalmente, estes valores foron comparados coas estimacións doutras poboacións de C. documentados hipopótamos na literatura. Este índice foi estimado da expresión:
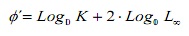
onde, k e l∞ son os parámetros de crecemento.
Dado que o valor estimado de L∞ neste estudo baseouse en LH e os documentados na literatura en lonxitude total (LT), a efectos de facer comparacións entre os valores, realizouse unha simple regresión lineal entre LT (dependente) e LH (independente) para facer a conversión de LH a lt. Para iso, o seguinte modelo lineal obtívose con datos deste estudo:
lt = – 0.4387 + (1.2335 lh)
Das frecuencias LH de capturas comerciais, a mortalidade total (Z), natural (m) e pesca (f) da poboación de C. Hippos procederase a estimar. Z estimouse a partir da curva de captura linealizada convertida a tamaños (17):
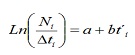
onde, nin é o número de individuos capturados pertencente á clase de lonxitude I; T’i é a era relativa da clase I; Δti é o tempo medio requirido polo individuo para o seu crecemento a través da clase i.
m estímase que utilizaba a ecuación empírica de Paully (17):
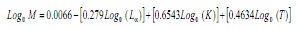
Onde, L∞ e K son os parámetros da ecuación de Von Bertalanffy e T é a temperatura media anual da auga costera da superficie na área de estudo, que foi considerada a 28,2 ° C.
A taxa de mortalidade de pesca e a taxa de operación estímase en:
Para realizar as estimacións dos parámetros, supuxéronse un crecemento estacional de C. hipopótamos, dado que ao longo do ano, episodios ambientais na área de estudo que poden estar influenciando o crecemento dos peixes. Tal é o caso das condicións estacionais do caudal do río Magdalena, que poden influír nas condicións físicas, químicas e biolóxicas na súa boca (8). A isto engádese, a influencia dos ventos de alisio no sector da boca de cinza, xa que cando están en marcha e segundo a súa intensidade, tamén cambian as condicións oceanográficas do sector (18). Do mesmo xeito, o coñecemento biolóxico das especies indica que C.Hipopótamos é unha especie migratoria (1), a partir da cal tamén se pode inferir a estacionalidade no crecemento. Baseado no anterior, os valores atopados por Elefan I foron seleccionados para estimar as taxas de mortalidade de C. Hippos neste estudo.
Co obxectivo de cuantificar o efecto das mostras mensuais nos parámetros de crecemento e Obtén os valores de precisión do mesmo, aplicouse a técnica de remuneración de Jackknife (6). Isto consiste en estimar os tamaños mensuais de frecuencia de tamaños e secuencialmente omitidos para obter combinacións de cada un dos parámetros. Para especificar esta estimación, utilizouse o erro porcentaxe (EP) eo coeficiente de variación (CV), utilizando as seguintes fórmulas:
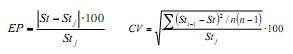
Onde, ST é o parámetro de crecemento estimado usando a base de datos anual total e STJ é o estimador medio derivado das sucesivas estimacións de Jackknife; STI-1 é o parámetro estimado ao omitir o mes I e n é o tamaño da mostra anual (n = 12).
Resultados
Durante o período de estudo, recolléronse 1151 individuos, dos cales 868 foron capturados con Transmal, 120 cun gancho e 167 con bolos; Presentando unha serie de tamaño entre 10,5 e 96,5 cm de lonxitude. Para a análise dos aspectos biolóxicos estudados, contáronse 264 individuos, sendo 180 machos e 84 mulleres, cunha variedade de tamaño entre 25 e 96,5 cm de LH e un peso total entre 281,5 e 10500 g.
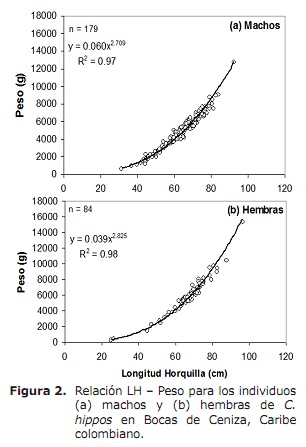
Relación de peso de lonxitude (LH-W). A análise revelou que entre o sexo a relación LH-W difería significativamente, observando que as femias pesan máis que os machos da mesma lonxitude (ANCOVA: F1, 260 = 21.5, P < 0.05 ). Polo tanto, a relación LH-W para os machos foi w = 0.060lh2.709 (p < 0.05) e para mulleres w = 0.039lh2.825 (P < 0.05) (Figura 2). Os valores de B resultantes das regresións para ambos sexos foron significativamente inferiores a 3, indicando un menor crecemento (T proba, P > 0.05; 15).
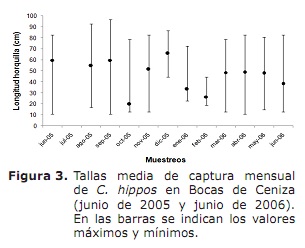
Composición de tamaño, tamaño medio da madurez sexual (L50%) e tamaño de captura media (TMC). En xeral, os tamaños mínimos de captura foron rexistrados en outubro, novembro de 2005 e marzo, abril e xuño de 2006; Aínda que as máximas eran frecuentes ao longo do período de estudo, excepto por outubro, xaneiro e febreiro (Figura 3).
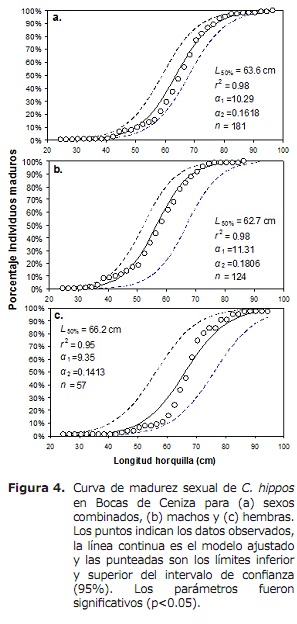
L50% para sexos combinados foi de 63,6 cm LH (95% CI: 59.0-68,4 cm). Para o caso dos machos e as femias foron de 62,6 cm (95% CI: 58.3-70 cm) e 66,2 cm (95% CI: 56.1-76,2 cm), respectivamente (Figura 4). O TMC global estimouse a 50,3 cm de LH. Para o caso de transmases, ganchos e bolos estímase en 56,0, 56,2 e 17,8 cm, respectivamente (Figura 5A, 5B e 5C), por baixo do L50%.
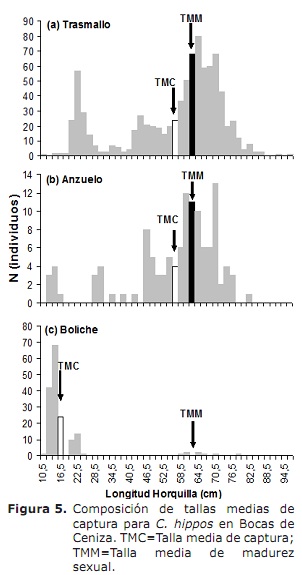
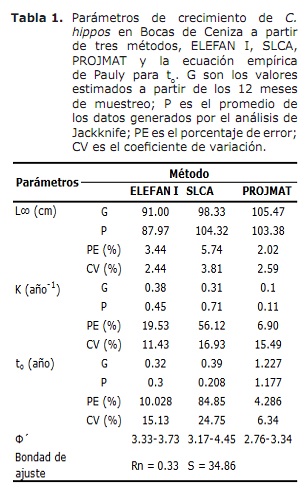
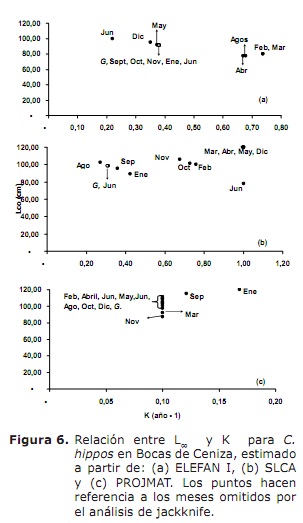
crecemento. Os resultados de L∞ e K proporcionados polos métodos Elefan I, SLCA e Projmat preséntanse na táboa 1. Observouse que estes tres métodos proporcionaron valores diferentes para cada un dos parámetros de crecemento. A análise de Jackknife mostrou que hai un efecto dos datos sobre os diferentes parámetros de crecemento, especialmente para SLCA e PROJMAT. Para o caso de Elefan I, L∞ varía de 77,4 a 100 cm, mentres que K valores estaban entre 0,20 e 0,75 anos-1 (Figura 6). Con SLCA, os valores máis altos de L∞ e K foron obtidos con valores entre 78,25 e 120 cm e entre 0,3 e 1,0 ano-1 respectivamente. Segundo os valores lanzados por Projmat, observouse menos dispersión para K fluctuando entre 0,10 e 0,17 anos-1. O contrario ocorreu por L∞, onde os valores variaron entre 87 e 120 cm, sendo o último máis grande que o observado nos estimadores de crecemento anteriores. Os valores Estimados K mostraron diferenzas entre os métodos, cos máis altos calculados por Elefan I e SLCA (Táboa 1). O valor obtido para a era hipotética á que o animal (a) comeza a crecer pola ecuación empírica de Paully (20) foi de 0,32 anos, cun coeficiente de variación do 13,92% e unha porcentaxe de erro (PE) de 10,02. A análise do índice de rendemento ǿ mostrou valores similares entre Elefan I (3.33-3.73) e SLCA (3.17-4.45), que non se produciron por projmat que mostraron valores máis baixos que os anteriores (2.76-3.34). Aínda así, a menor variación foi presentada para Elefan I, seguida de Projmat e SLCA, sendo este último o rango máis alto presentado. Tendo en conta o coeficiente de variación (CV) de cada un dos parámetros, descubriuse que o Elefan I estima que a análise de Jackknife tendía a proporcionar valores máis baixos para L∞, a diferenza de SLCA e Projmat que alcanzaron valores máis altos (Táboa 1) ..O resultado do PE obtido para K con projmat foi menos do que se atopou para Elefan I e SLCA. En consecuencia, o método que arroxou as estimacións máis credíveis dos parámetros de crecemento foi Projmat, seguido por Elefan I e finalmente SLCA.
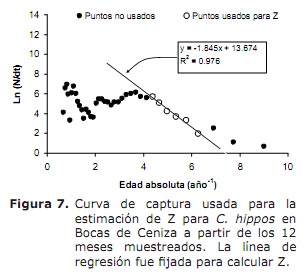
Estado de mortalidade e explotación. Baseado nos seguintes parámetros de crecemento: k = 0,38 ano-1; L∞ = 91 cm LH; a = 0,32 anos; Oscilación estacional no patrón de crecemento (c) de 0,5 e unha taxa de crecemento mínimo (WP) de 0,3, unha taxa de mortalidade total (Z) de 1,76 ano-1 (figura 7) foi estimada, oscilando entre 1.38 e 2,22 anos -1 ( da análise de Jackknife).
A mortalidade natural (M) foi de 1,12 ano-1, variando entre 1,10 e 1,14 anos-1. A taxa de mortalidade pesqueira (F) estímase en 0,63 anos-1, presentando unha variedade de variación entre 0,2 e 1,06 anos-1. Os valores da proporción F e Z para estimar a taxa operativa (e), presentaron un valor máximo de 0,5 anos-1, unha media de 0,30 anos-1.
Discusión
Relación de peso de lonxitude (LH-W). O concepto de crecemento implica cambios relativos no tamaño e peso dos individuos (19), ea súa relación ten varios usos na investigación pesqueira. Por exemplo, é útil nos modelos de análise de Stock Pesca para estimar a biomasa dunha poboación; Coñecer a condición dun peixe; Comparar a historia da vida de certas especies entre rexións; e para determinar o tipo de crecemento que o peixe normalmente presente (20). C. hipopótamos, presenta un peso sen peso de tipo alométrico (< 3; 15), indicando que presenta taxas de crecemento diferencial para diferentes partes do corpo, onde a lonxitude aumenta a A taxa máis alta que o peso.
Os resultados indican que esta condición foi máis evidente para os homes que para as mulleres, xa que este último, a pesar de que presenten un menor crecemento alométrico, aproximado para ter crecemento isométrico. Isto foi concordante co resultado de ANCOVA, o que indicou que o mesmo LH hai diferenzas de peso sexual. As femias desta especie presentan maior peso e altura corporal que os machos. Doutra banda, é importante destacar que porque as mostras foron recollidas durante un ano, a relación LH-W neste estudo non representa unha época particular do ano. Neste sentido, os valores B poderían considerarse como un valor medio anual para a especie (20). Os resultados desta investigación foron similares aos documentados para a especie en diferentes áreas xeográficas. Por exemplo, os valores B baseados en B foron documentados a partir de 2.73 en South Florida (21), a partir de 2,85 na India (1) e 2.91 no Golfo de Salamanca, Colombia (22).
Crecemento, Estado de mortalidade e explotación. Con base nos parámetros de crecemento obtidos por Elefan I, a lonxitude máxima estimada para C. Hippos en Ash Bocas foi de 91 cm LH correspondente a 111,81 cm de lt. Se este último se converte en idade correspondente a 14 anos, alcanzado cunha taxa de crecemento K de 0,38 anos-1, o que suxire que C. Hippos é unha especie de longeve (17). Doutra banda, tendo en conta a curva de crecemento, esta especie creceu rapidamente ata a idade de 5 anos. Este dato coincide con outras presentacións, como a de Trachinotus Falcatus, que alcanza un tamaño máximo de 90 cm e tamén ten un crecemento máximo de 5 anos, desde o que inicia a fase estacionaria do seu crecemento (2).
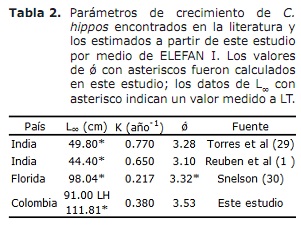
Por outra banda, os valores de L∞ estímase que C. hipopótamos neste estudo foron superiores aos documentados na literatura; Mentres os do índice de rendemento ǿ revisados na literatura mostraron valores moi próximos á estimación neste estudo (Táboa 2). Tomando este índice como criterio de comparación entre os valores dos parámetros de crecemento obtidos e tendo en conta que C. Hippos é unha especie migratoria (23), pódese supoñer que o crecemento desta especie é concordante que se describe en diferentes áreas da súa distribución global (Táboa 2).
O patrón de crecemento anual de C. HIPPOS mostrou oscilacións (c = 0,5) cunha taxa de crecemento mínimo no mes de setembro (WP = 0.3). Este patrón pode estar influenciado por un dos aspectos máis característicos do crecemento dos peixes: a variabilidade observada entre individuos dunha poboación e entre as poboacións dunha especie, que está directamente relacionada co medio ambiente (24). Por exemplo, a variabilidade das condicións físico-químicas da auga (24) ea calidade e cantidade de alimentos (19) condicionan a migración dunha especie, que engloba as desprazamentos da estación entre as zonas de reprodución, alimentación e desova (15, 25) ..
Con base no anterior e tendo en conta as frecuencias de lonxitudes observadas durante o período de estudo, que tamén indican a posible presenza de estacionalidade no crecemento (13), pódese inferir que o crecemento de C. Hippos é estacional. En canto á taxa de crecemento mínima, este evento pode ser impartido pola etapa reprodutiva da especie, xa que ata setembro observouse un aumento no número de individuos maduros (en etapas III).
Por outra banda, a relación entre a taxa de mortalidade de pesca (f) ea mortalidade total (Z) de 0,3, indica que a poboación na área de boca de cinza non está correndo risco de ser sobreexplotación , baixo a premisa de que unha poboación alcanza a súa actuación óptima cando a fracción de morte causada por F é superior a 0,5 (6,13). Non obstante, é importante resaltar que cando o L50% foi comparado con TMC, C. Hippos presentou unha maior presión sobre a poboación adulta, frecuentemente capturada con Transmal e Hook. A situación empeora co uso de Boliche, que non só afecta aos adultos, senón tamén aos mozos, que non permiten a renovación natural deste recurso de pesca.
Proposta para o deseño dun sistema de manexo de pesca .. A partir da base que a sobrepesca pode ter lugar en tres xeitos diferentes: (i) en relación ao crecemento; (ii) con respecto ao reclutamento; e (III) Overpishing que afecta ao ecosistema (15). Neste sentido, a pesca de C. hipopótamos no sector da boca de cinza pode clasificarse dentro do primeiro elemento, que ten lugar cando os individuos son capturados antes de alcanzar o tamaño necesario para garantir a renovación biolóxica da poboación. O anterior tornouse un problema común para a maioría das especies comerciais, e foi documentado para moitas especies costeiras no Caribe colombiano.
Todos os anteriores, é necesario que os esforzos están investindo para proxectar estratexias de xestión para C. Hipopótamos na pesca de Asho Bocks, de tal xeito que son consistentes co enfoque de precaución á pesca responsable deste recurso (27). Caddy (27) suxire que as estratexias deben aplicarse dentro dun marco de redundancia de xestión onde tal marco podería consistir nunha mestura de medidas que impliquen artes, criterios secantes temporais e o aumento dos tamaños mínimos de captura. Neste sentido, tendo en conta o L50% como punto de referencia límite (28), aquí recoméndase que o tamaño mínimo de captura deste recurso sexa fixado a 64 cm de lonxitude. Do mesmo xeito, en todos os engrenaxes de pesca utilizados, proponse aumentar os tamaños de malla superior a 8,89 cm e usar calibres de gancho destinados a capturar peixes máis grandes. Nas condicións actuais, estes equipos de pesca afectan directamente a fracción de mozos que entran na primeira fase de madurez sexual. Coas medidas anteriores, complementadas con outras variables como a captura, o esforzo de pesca e a captura por unidade de esforzo, así como os factores sociais e económicos, espérase que C. Hippos sexa capturado nunha maior proporción de individuos superiores ao L50% Para garantir a sostibilidade deste recurso ao longo do tempo. Neste sentido, é importante ter en conta o estado multispecífico e multiflot da pesca e, polo tanto, recoméndase un proceso de acordo entre os pescadores, os comerciantes e as entidades relevantes para chegar a acordos dirixidos a unha explotación responsable do recurso.
En conclusión, o Jorel non presenta a sobreexplotación en boca de cinza. Non obstante, ao comparar o tamaño medio de captura co L50%, a especie está sendo capturada cos principais artes de pesca antes do 50% da poboación consegue madura. As directrices para o deseño dun sistema de pesca que garante a conservación e uso sostible deste recurso son propostas.
Recoñecementos
á comunidade de pescadores, os membros de Asropscar e Coopez. Para os irmáns Villa e a Pescadería o Nuevo tambó pola súa colaboración na coordinación de actividades e subministración de información de campo. Para os profesores G. Lozano, L.C. Gutiérrez, O. Galvis, J. Branco e E. VILORIA.
Referencias
1.REUBEN, S, HM KASIM, S SIVAKAMI, PN Radhakrihnan, KN KURUP, M Sivila, Nobre, KVS Nair, Raje SG. Pesca, bioloxía e avaliación de accións dos recursos carangid dos mares indios. Indian J Fish 1992; 39 (3,4): 195-234.
2.Crabtree, R, PB Hood, Snodgrass D. Idade, Grouth e Reprodución do permiso (Trachinotus Falcatus) en Florida Waters. Peixes de 2002; 100: 26-34.
3.river R, SOLANO, G.Diagnóstico de actividade pesqueira artesanal na zona costeira do Departamento Atlántico, Caribe Colombiano. Barranquilla, Colombia: Incodador; 2005.
4.Gayalino, FC, P Sparre, Paully D. A guía do usuario Fisat. FAOL – ferramentas de asiso de Stock Iclarm. Roma: FAO; 1993.
5.Sparre, P., VENEMA S. Introdución á avaliación dos recursos pesqueiros tropicais. Roma: FAO; 1995. Documento de pesca técnica: 306/1.
6.Rueda, M, Santos-Martínez A. Dinámica da poboación da laya Mujarra Eugerres Plumieri da Big Ciénaga de Santa Marta, Colombia. Peixe RES de 1999; 42: 155-166.
7.Blanco, JA, EA VILORIA, Narváez BJC. Enso e cambios de salinidade interunnual no Sistema de Lagoa Ciénaga de Santa Marta, Caribe Colombiano. Estuar a costa shelf sci 2006; 66: 157-167.
8. RESTREPO, JD, KJERFVE B. Río Magdalena: variabilidade interunnual (1975-1995) e descarga de auga revisada e estimacións de carga de sedimentos. J Hydrol 2000; 235 (1-2): 37-149.
9.AveVedo a, gutiérrez ml. Algúns aspectos reprodutivos do riztrionodón poroso Toyo (1831) (Elasmobranchii: Carccharinidae). No sector da boca de cinza, o Caribe Colombiano. .. Barranquilla, Colombia: Universidade Atlántica, Facultade de Ciencias Básicas; 1999.
10.Vazzoleler, EAM. Método Manual para Peixes Populare Estudantes biolóxicos: reprodución e crescimento. Brasília: CNPQ; 1982.
11.Sheperd, jg. Un método débilmente paramétrico para a análise de datos de composición de lonxitude. En: Paully, D, Morgan GP editores. Métodos baseados en lonxitude na investigación pesqueira. Manila, procedementos de conferencia de ICLARM 13. 1987.
12.Lielie, pH. O uso de matrices en determinadas matemáticas de poboación. Biometrika 1945; 35: 213-245.
13.Gulland, JA, ROSEBERG AA. Exame dos métodos que se basean no tamaño para avaliar as poboacións de peixes. Roma: FAO; 1992.
14.Tordecilla-Petro G, S Sánchez-Band, Olaya-Nieto Ch. Crecemento e mortalidade de Moncholo (Hoplias Malabaricus) na Lorica Lorica, Colombia. REV MVZ CÓRDOBA 2005; 10 (2): 623-632.
15.Pauly, D. Algúns métodos sinxelos para a avaliación dos recursos pesqueiros tropicais. Roma: FAO; 1983.
16.Pauly, D, Munro JL. Once máis sobre a comparación de crecemento en peixes e invertebrados. Fishb 1984; 1:21
17.Pauly, D. Dinámica de poboación de peixes en augas tropicais; Un manual de uso con calculadoras programables. IClom Stud Rev 1984; 8: 235.
18.blanco, JA. Variacións ambientais estacionais nas augas costeiras ea súa importancia para a pesca no Santa Marta, a rexión do Caribe Colombiano. .. Santa Marta-Bogotá. Universidade Nacional de Colombia, Facultade de Ciencias Básicas; 1988.
19.Abaunza L, Karlou-Riga C, Murta A, Eltink W, García Santamaría MT, Zimmermman C, Hammer C, et al. Crecemento e reprodución de cabalos de cabalo, trachurus trachurus (carangidae). Rev Fish Biol Fish 2003; 13: 27-61.
20.Froese, lei de R. Cube, factor de condición e relación de peso: historia, meta-análise e recomendacións. J APPLAY ICHTHYOL 2006; 22: 241-253.
21.BOHSACK, JA, Harper de. Relacións de lonxitude de peso dos peixes de arrecifes mariños seleccionados do sueste dos Estados Unidos e do Caribe. Memorando técnico de NOAA; 1988. NMFS-SEFC-215.
22.duarte, Lo, CB García, Moreno, G Melo, P Navajas, N Sandoval, et al. Atlas demográfico do Golfo de Salamanca, Caribe Colombiano: dinámica de poboación, distribución, comida e reprodución. .. Versión 1. Santa Marta: Invemar-Unal-Colciencias; 1999.
23.HoneyBrink, R. Unha revisión da bioloxía da familia Carangidae Whith enfassis sobre as especies atopadas nas augas hawaianas. Honolulu: División de Recursos Acuáticos; 2000.
24.Macer, CT. Algúns aspectos da bioloxía do cabalo de cabalo (Traquurus trachurus (L)) en augas en torno a Gran Bretaña. J Fish Biol 1977; 10: 51-62.
25.Harden, fr. Migración de peixes. Londres: Arnold; 1968.
26.Narváez B, JC, F Herrera, J. Efectos J. Efectos da pesca sobre o tamaño do peixe nunha pesca artesanal caribeña colombiana. Bol Invest Mar custo 2008; 37 (2): 163-187.
27.caddy, jf. Xestión de pesca no século XXI: aplicaranse novos paradigmas?. Rev Fish Biol Fish 1999; 9: 1-43.
28.caddy, jf. Limitar puntos de referencia, semáforos e enfoques holísticos á xestión da pesca con entrada mínima de avaliación de accións. Peixe RES 2002; 56 (2): 133-137.
29.Torres, FJR, MBTM NORIZAM, LR GARCÉS, SILVESTRE GT. Compilación de parámetros de poboación de peixes especies comúnmente capturadas en arrastre en sur e sueste asiático. Manila: World Fish Center; 2001.
30.Snelson, ff. Estudos biolóxicos de Crevalle Jack (Caranx Hippos) en Florida. St. Petesburg: Florida Marine Reseach Institute; 1992.