Aliments funcionals
L’expressió d’IL-10 intervé en la regulació de la resposta inflamatòria pels àcids grassos omega 3
S. Serra, F. Lara-Villoslada, el Sr. Olivares, J. Jiménez, J. Boza i J. Xaus
Departament d’Immunologia i estudis preclínics. Puleva Biotech. Granada.
|
Resum Objectiu o antecedent: Els àcids grassos poliinsaturats són importants per a l’organisme humà a causa de la seva implicació en nombroses funcions biològiques. Les dietes occidentals es caracteritzen per ser excessivament riques en àcids grassos w-6 i pobres en àcids grassos w-3. : Els àcids grassos de la sèrie w-3 són necessaris per al normal creixement i desenvolupament de l’individu i per a la regulació de la resposta immunològica. L’objectiu d’aquest estudi és analitzar l’efecte d’una dieta enriquida en àcids grassos w-3 davant d’un procés inflamatori així com l’estudi dels mecanismes implicats en aquest efecte. (Nutr Hosp 2004, 19: 376-382) Paraules clau: àcids grassos poliinsaturats. Inflamació. Organisme. |
IL-10 EXPRESSION IS Involved IN THE REGULATION oF THE immune RESPONSE BY OMEGA 3 fatty acids Abstract Introduction: poliinsaturats fatty àcids play a key role in a huge number of biological functions. Western Diets are highly rich in w-6 fatty acids. However the content of w-3 fatty àcids is not suitable in those Diets, despite of their importance in normal development of the human body and regulation of immune response. (Nutr Hosp 2004, 19: 376-382) Key words: poliinsaturats fatty acids. Antiinflammatory. Organism. |
Correspondència: J. Xaus. del Departament d’Immunologia i estudis preclínics. a Puleva Biotech. del Camí de Purchil, 66. 18004 Granada.
Correu electrònic: [email protected]
Rebut: 21-I-2004.
Acceptat: 13-II-2004.
Introducció
El sistema immunològic està involucrat en la defensa de l’organisme enfront d’agents estranys, cèl·lules tumorals i agressions externes. Un elevat nombre d’òrgans, cèl·lules, teixits i molècules interactuen per dur a terme aquesta missió.
El desenvolupament, manteniment i òptim funcionament de sistema immunològic depèn de la dieta entre altres factors. Entre els nutrients que més afecten el sistema immunològic destaca el greix la deficiència o desequilibrada aportació així com la qualitat de la mateixa comprometen l’estabilitat de sistema inmunológico1.
La relació entre àcids greixos i inflamació es deu principalment a l’component fosfolipídico de les cèl·lules de el sistema immunològic. Els àcids grassos incorporats a la membrana de les mateixes poden afectar les funcions d’aquesta i per tant a l’funcionament cel·lular. Els fosfolípids de membrana són responsables de la fluïdesa de la mateixa, que afectarà els diferents components ancorats en ella. Diverses senyals intracel·lulars parteixen d’aquests fosfolípids i la seva composició, per tant, afecta diferents vies de transducció de senyals. A més, els àcids grassos sintetitzats a partir dels fosfolípids, són substrats per a la síntesi d’un grup de mediadors, els eicosanoides, de marcada importància biológica2.
Actualment, l’interès per l’estudi dels àcids grassos és creixent, i cobra importància la influència d’ells sobre la resposta immunològica. Aquest interès s’ha centrat en l’estudi dels àcids grassos poliinsaturats de cadena llarga (AGPI-CL), procedents de les famílies w-6 i w-3, els precursors, àcids linoleic i α-linolènic, són essencials, havent de ser ingerits amb la dieta, així com es recomana també incloure-hi alguns dels seus derivats, com l’àcid eicosapentaenoic (EPA), docosahexaenoic (DHA) i araquidònic (AA), ja que a través de les vies metabòliques, per elongació dels seus precursors, no s’assoleixen els nivells adecuados3.
En individus que consumeixen dietes occidentals, les cèl·lules inflamatòries contenen elevades proporcions d’AGPI-CL de la sèrie w-6, especialment d’AA i baixes proporcions, però, de l’EPA de la sèrie w-34. El principal paper d’AA és actuar com a substrat per a la síntesi de mediadors bioactius de l’tipus eicosanoides. Aquests estan implicats en modular la durada i la intensitat de la resposta inflamatòria. Els derivats de l’AA, com TXA2 i LTB4, són potents agents amb un marcat efecte infamatorio. El TXA2, té efecte vasoconstrictor i proagregante mentre que el LTB4 és un potent agent quimiotáctico5. A més, la PGE2 indueix febre, augmenta la permeabilitat vascular i la vasodilatació, a el temps que la producció d’IgE per cèl·lules B.
L’augment de l’consum dels àcids grassos w-3 comporta una disminució de l’AA en les membranes de les cèl·lules inflamatòries disminuint-així la seva capacitat per sintetitzar a partir de la COX i la LOX eicosanoides derivats de l’AA. A més d’inhibir el metabolisme de l’AA, l’EPA inhibeix l’oxigenació de l’AA per la COX, i suprimeix la inducció de certes citocines que actuen sobre l’expressió gènica de la COX-2 i la 5-LOX. També és capaç d’actuar com a substrat de la COX i la 5-LOX, donant lloc a eicosanoides considerats biològicament menys actius5.
Tot això porta a la idea que l’oli de peix té propietats antiinflamatòries. El consum en individus sans d’una proporció adequada dels àcids grassos w-6 i w-3 portaria a una millor regulació de la resposta immunològica, i el consegüent increment de la proporció dels àcids grassos w-3 resultaria beneficiós per a subjectes que pateixin malalties autoimmunes o inflamatòries per l’efecte antiinflamatori dels mismos1.
No només aquests mecanismes estan involucrats en l’efecte antiinflamatori sinó que actualment s’està coneixent la seva implicació a nivell de vies de senyalització com NFkB, MAPK o PPAR i LXR6. L’actuació a través d’aquests dos últims receptors obre un nou camp d’investigació i una major gamma d’aplicacions dels àcids grassos w-3 a causa de la localització i funció dels mateixos.
El present estudi tracta de posar de manifest l’efecte antiinflamatori dels àcids grassos w-3 i els mecanismes implicats, mesurant paràmetres immunològics que permetin comprovar el seu efecte a nivell sistèmic, i analitzant el curs d’una inflamació induïda per determinar la seva actuació a nivell local.
Material i mètodes
Reactius i anficuerpos
La concanavalina A (Amb A) i el lipopolisacàrid (LPS) van ser proporcionats per Sigma Chemical Co (St Louis, Mo). Per mesurar immunoglobulines es van utilitzar anticossos proporcionats per Bethyl (Montgomery TX) i en el cas de les citocines per Biosource (Camarillo, CA). La peroxidasa de rave, el bromur de hexadeciltrimetilamonio, el clorhidrat d’o-dianisidina i el peròxid d’hidrogen per a la mesura d’activitat miloperoxidasa així com l’àcid tricloroacètic, el ß-NADPA, el 5,5-ditiocarbámic-bis (2-àcid nitrobenzoico) i el glutatió van ser també adquirits en Sigma. La glutatió reductasa va ser subministrada per Roche.Es va emprar aigua desionitzada purificada amb un sistema Millipore Milli-Q (Bedford, MA).
Animals i dietes
Ratolins mascle Balb / c a l’deslletament de tres setmanes d’edat van ser proporcionats per la Universitat de Granada. Se’ls va mantenir durant tot l’experiment a una temperatura controlada de 22 ° C i sota un cicle de llum de 12 hores. L’experiment global va ser dut a terme 3 vegades amb resultats similars.
Tots els estudis animals que es van realitzar es van dur a terme segons les Normes d’Ètica Animal fixades en els Criteris d’Hèlsinki.
l’experiment consta de 2 grups de ratolins (n = 10) alimentats durant 28 dies amb dietes base iguals i la diferència rau en el tipus de greix inclosa. Totes dues contenen el 5% de greix de l’total de la dieta. Aquest 5% de greix està constituït per oli de gira-sol en el cas de el grup usat com a control. En el cas de la dieta w-3, el 12% de l’contingut total de greix és oli de peix refinat, mentre que la resta del greix és el mateix oli de gira-sol de el grup anterior. Ambdues dietes van ser elaborades de mane-ra específica seguint les recomanacions de l’Acadèmia Americana de Veterinària per Nutrició Animal. D’aquesta manera es van preparar dietes semisintètiques incloent-hi el 5% de greix, 21,3% de proteïna, 67,5% hidrats de carboni, 5,2% de fibra i l’1% restant d’una barreja de vitamines i minerals.
Les dietes van ser conservades en atmosfera d’argó i administrades diàriament als ratolins per evitar l’oxidació dels AGPI-CL. Al llarg de l’estudi es va dur a terme un control de l’pes i aspecte.
Dotze hores abans de l’sacrifici es va induir una dermatitis de contacte en una de les orelles de cada ratolí. L’agent irritant usat va ser el 2,4-dinitro-1-fluorobenzeno a l’0,5% en una solució d’acetona: oli d’oliva 4: 1. Es van utilitzar d’aquesta solució 80 ml que es van posar en cada oreja7.
El sacrifici es va realitzar per mitjà d’una injecció intraperitoneal de pentotal sòdic (50 mg / kg) i seguidament la sang va ser extreta per punció cardíaca en tubs que contenien EDTA (60 mg / ml). La sang es va centrifugar (3.500 rpm) 10 minuts a 4 ° C i les alíquotes de sèrum van ser recollides i emmagatzemades a -80 ° C.
Processament de les orelles
Les dues orelles de cada ratolí van ser pesades en balança de precisió. La meitat d’elles van ser introduïdes durant 24 hores en una estufa a 60 ° C i posterior-ment es va analitzar el seu pes sec i el seu consegüent contingut d’aigua. La meitat restant, es va homogeneïtzar independentment en tampó HITAB (Bromur de hexadeciltrimetilamonio) a la proporció de 1/20 per a procedir a la mida de glutatió total i d’activitat mieloperoxidasa. Aquestes mesures es van realitzar segons el descrit anteriormente8,9.
Cultiu de cèl·lules
Els melses de cada ratolí es van extreure i van conrear com s’ha descrit anteriormente10. La medul·la òssia dels fèmur i les tíbies de cada ratolí van ser extrets per injecció de mitjà i conreades, prèvia diferenciació, segons s’ha descrit amb anterioridad1l.
Assaig de proliferació
Els limfòcits derivats de melsa van ser cultivats en plaques de 24 pous (2 x 106, cèl·lules / pou) amb 500 ul de mig i incubats en presència o absència de amb a (5 mg / ml) o LPS (50 mg / ml). Timidita tritiada (Amersbam Biosciences, Arlington Heights, IL) a la concentració d’1 μCi / ml va ser afegida a cada pou. La incorporació de timidina després de 48 hores va ser analitzada com s’ha descrit anteriormente1l. Les mesures es van realitzar per duplicat i van ser representades com la mitjana ± SD.
Producció de citocines
Els limfòcits de melsa van ser cultivats en plaques de 6 pous (5 x 106 cèl·lules / ml ) amb 5 ml de medi i van ser estimulats amb amb a i LPS a les mateixes concentracions usades en l’assaig de proliferació. Després de 48 hores es van recollir els sobrenedants. El sediment va ser lisat amb 1 ml de trizol per a posterior extracció de RNA. En el cas dels macròfags, aquests van ser cultivats en plaques de 6 pous (3 x L06 cèl·lules / pou) i estimulats amb LPS (100 ng / ml). Després de 24 hores es va recollir el sobrenedant i es lisaron els sediments cel·lulars.
Els nivells d’IgA, IgE, i IgG1, van ser determinats en sèrum. La concentració de IL-10, TNFα, i IL-2 va ser mesurada en sobrenedant de melsa i de macrófiagos per ELISA seguint les instruccions de fabricant.
Anàlisi estadística
El grau de significació estadística entre els grups va ser determinat pel test de la t-Student a dues cues i amb variàncies iguals entre les mostres (p < 0,05). Els resultats han estat expressats com la mitjana ± SD.
Resultats
Els animals van ser controlats durant el mes que va durar l’experiment observant una conducta i aspecte normals. Es va comprovar que l’evolució en el creixement era l’adequada i que no hi havia diferències entre els dos grups.No va existir tampoc cap tipus d’anomalies pel que fa a la quantitat, consistència i aspecte de la femta, sent el contingut d’aigua de les mateixes similar en ambdós grups.
Amb l’objectiu d’avaluar els efectes dels àcids grassos w-3 en els rosegadors es van estudiar diversos paràmetres sistèmics que poguessin aportar informació de com aquests modulen el sistema immunològic i per tant, aclarir els seus possibles mecanismes d’acció.
Les citocines són un grup molt divers de molècules produïdes per cèl·lules en resposta a una gran varietat d’estímuls i que són capaços d’alterar d’alguna manera el comportament d’altres cèl·lules. És el cas de la IL-10, un interleucina de tipus regulador que hi ha l’anomenada resposta Th3. En aquest estudi es van prendre sèrum, sobrenedant de cultiu de limfòcits i sobrenedant de macròfags i es va determinar per ELISA els nivells d’aquest mediador, descubrién-se un augment significatiu en el grup de ratolins alimentats amb àcids grassos w-3 a la producció d’IL -10 per part de macròfags (94,7 ± 57,8 vs 126,5 ± 37,6 pg / ml, p = 0,05) i limfòcits (31,56 ± 5,51 vs 61,29 ± 29,08 pg / ml p = 0,045) a l’ésser les cèl·lules estimulades (fig. l).

Fig. 1.-El consum d’àcids grassos w-3 indueix l’expressió a d’IL-10. Nivells d’IL-10 en sobrenedant de cultiu de melsa
sense estimular o estimulat amb concanavalina A (Amb A).
a Com a conseqüència de l’increment d’IL-10 es va observar la disminució de citocines de tipus Th1, com ara la IL-2 alliberada per limfòcits (332,83 ± 197,86 vs 181,61 ± 137,1 pg / ml, p = 0.05) (fig. 2A). El descens en els nivells d’aquesta interleucina va portar a la disminució significativa de la proliferació de limfòcits B en el grup de ratolins tractats amb w-3 (7.300 ± 2.873,55 vs 4.681 ± 2.243,35 cpm, p = 0,04) ja una reducció de la proliferació dels limfòcits T (7.622,6 ± 3.692,8 vs 5.048,4 ± 2.507,07 cpm, p = 0,07) assenyalant ia un possible efecte supressor dels àcids grassos w-3 sobre el sistema immunològic (fig. 2B). Aquesta dada es va correlacionar també amb una tendència a disminuir la mida de la melsa expressat com mg melsa / g ratolí (5,72 ± 1,36 vs 4,7 ± 0,54, p = 0,08).

Aquesta disminució en el nombre de limfòcits ha de tenir una repercussió dràstica sobre el subjecte causa de el paper fonamental d’aquestes cèl·lules en la defensa de l’organisme. Per tant, altres citocines alliberades per aquestes cèl·lules podrien estar alterades. També l’efecte de la IL-10 hauria d’haver recaigut sobre els macròfags de manera que per comprovar si es produïa aquesta correlació es va procedir a mesurar alguns d’aquests mitjana-dors com, per exemple, el TNFα (5.306 ± 0,51 vs 3.831 ± 0,82 ng / ml, p = 0,06) (fig. 2C).
A més de la disminució de la resposta tipus Th1, també observem que la dieta rica en w-3 exercia un efecte inhibidor de la resposta Th2 com s’observa per la disminució dels nivells plasmàtics d’IgE (291, 7 ± 53,06 vs 176,04 ± 71,4 ng / ml, p = 0,06) i IgG (802,31 ± 228,1 vs 392,01 ± 191,3 mg / ml, p = 0,05 ) (fig. 3A, B) o d’IL-4 i histamina (dades no mostrades).

a tot i la disminució generalitzada de la resposta immunològica, els ratolins alimentats amb w-3 no van experimentar símptomes de debilitat ni predisposició a la infecció. Això pot ser degut a l’increment en aquests animals de paràmetres relacionats amb la defensa natural de l’organisme com poden ser els nivells d’IgA sèrics (208,23 ± 100,3 -vs 572,26 ± 135,01 mg / ml, p = 0 , 04) (fig. 3C), expressió de iNOS i la producció de nO pels macròfags (dades no mostrades).
Tots aquests resultats suggereixen un paper antiinflamatori dels àcids grassos w-3. Per reafirmar aquest paper es va emprar en aquests mateixos ratolins un model de dermatitis de contacte on intervenen tant la resposta Th1 com la Th2. Es va dur a terme a les orelles, i l’anàlisi de les mateixes va aportar informació sobre el curs de la inflamació i sobre les conseqüències palpables de l’administració d’àcids grassos w-3.
Després de la inducció de la dermatitis de contacte, les orelles van ser tallades i ràpidament es van analitzar els seus pesos, observant-se un increment més gran de pes a causa de la formació de l’edema, pel que fa a la sana, en els ratolins les dietes contenien oli de gira-sol només (168,041 ± 20,16 vs 113,024 ± 11,9 expressat en%, p = 0,0003) (fig. 4A); el seu pes sec es va determinar veient que en ambdós grups s’adquirien valors semblants i que, per tant, l’increment de pes experimentat, és exclusivament a causa de l’extravasació de líquids i a la migració de cèl·lules cap al focus inflamatori. Es va avaluar el contingut de líquid de les orelles, comprovant-se que també era molt superior en el grup de ratolins alimentats amb àcids grassos w-6 (19,4 ± 4,5 vs 3,94 ± 2,92 mg, p = 0,002) (fig. 4B).
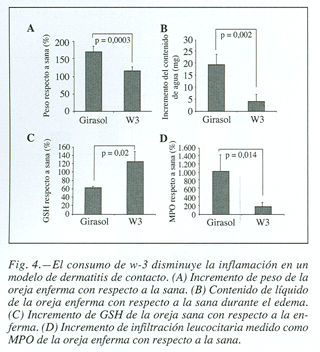
a Es van determinar altres paràmetres que aportessin més dades sobre el concurs de l’procés inflamatori, En aquest sentit, el glutatió de les orelles dels ratolins que van consumir dietes riques en àcids grassos w-3, era significativament superior a el de el grup control (61,04 ± 4,5 vs 123,69 ± 24,2 expressat en%, p = 0,02) (fig. 4C).
la major gravetat de la inflamació també va ser corroborada amb la mesura de l’activitat mieloperoxidasa. Aquest enzim està present en els grànuls dels neutròfils i catalitza la producció de HClO, constituint un marcador sensible d’infiltració de neutròfils i d’inflamació. Es va observar una major infiltració leucocilaria a les orelles inflamades de ratolins que havien consumit oli de gira-sol (163,19 ± 52,9 vs 1.009,74 ± 404,69 expressat en%, p = 0,014) (fig. 4D).
Tots aquests resultats indiquen que la incorporació a la dieta dels àcids grassos w-3 produeix una regulació de el sistema immunològic que fa que l’individu estigui menys predisposat a patir processos inflamatoris. La modificació d’un gran nombre de paràmetres sistèmics entre els quals destaca l’increment de la IL-10, podrien ser els responsables de la protecció exercida pels àcids grassos w-3 de la dieta.
Discussió
Entre els àcids grassos, els de la sèrie w-3 semblen ser els de major activitat immunomoduladora i dins dels w-3, són els derivats de l’peix, EPA i DHA, els biològicament més activos4. Nombrosos experiments en animals i estudis d’intervenció clínica indiquen que els àcids grassos w-3 tenen propietats antiinflamatòries i que per tant podrien ser útils en el tractament de malalties inflamatòries, i autoimmunes.
La primera evidència de la importància d’incloure àcids grassos w-3 en la dieta a nivell d’inflamació, va sorgir per observacions epidemiològiques de la baixa incidència de malalties autoimmunes i inflamatòries en els esquimals de Groenlàndia. Poc més tard es produïa un període d’expansió en el coneixement dels PUFA en general, i dels w-3 a particular4.
Actualment se sap que els àcids grassos w-3 són essencials per al normal creixement i desenvolupament de l’individu i que juguen un paper clau en la prevenció i tractament de malalties cardiovasculars, hipertensió o artritis entre altres4.
Malauradament, en les dietes occidentals predomina el consum en excés d’àcids grassos w-6, metabòlicament diferents als w-3 i amb funcions fisiològiques generalment oposades. Aquest desequilibri contribueix a l’augment en la incidència de problemes cardiovasculars i desordres inflamatorios4. Per tot això, esdevé un repte per a nombrosos científics, el demostrar i donar explicació a l’efecte antiinflamatori dels àcids grassos w-3.
Els nostres resultats suggereixen que els àcids grassos w-3 disminueixen la proliferació de limfòcits probablement a causa de l’efecte que exerceixen sobre la producció d’IL-2 i altres citocines implicades en la proliferació i diferenciació limfocitària. Aquests mateixos resultats han estat trobats per nombrosos estudis en animals de experimentación12,13. En estudis clínics en humans on es van incloure àcids grassos w-3 també s’han analitzat aquests paràmetres, trobant disminució de proliferació de limfòcits, encara que en aquest cas sense afectar-se els nivells d’IL-214. No obstant això, els resultats aportats pels estudis en humans són conflictius, trobant-se alguns treballs, fins i tot d’ell mateix grup, on no es modifica15 o fins es aumenta16 la proliferació de limfòcits. Aquestes diferències poden ser degudes a: Les dosis d’àcids grassos provats en estudis humans no es poden comparar amb les elevades dosis administrades en animals; els preparats d’oli de peix que s’utilitzen en els diferents estudis varien seves concentracions d’EPA i DHA pot variar també els efectes, i finalment la majoria dels estudis humans no han estat prou consistents per tenir en compte la gran variació en alguns paràmetres de la resposta immunològica.
Els àcids grassos w-3, a més de disminuir la producció de la citocina IL-2, disminueix una altra sèrie de citocines també Th1, com són la IL-1ß i el TNFα produïdes per macròfags. En humans sans, els nivells d’ambdues citocines també són regulats pels àcids grassos w-317 suggerint que l’efecte antiinflamatori podria estar mediat per la modulació exercida sobre aquestes citocines.
Pel que fa a la resposta Th2, el nostre estudi demostra també una disminució de la mateixa ja que hem observat una reducció dels nivells sèrics d’histamina, IgE i lgG1, així com de citocines Th2 com la IL-4. Hi ha diverses evidències epidemiològiques que donen suport al paper protector dels àcids grassos w-3 en malalties de tipus al·lèrgic.No obstant això, estudis usant suplements de peix en asma revelen un escàs impacte clínic tot i els canvis bioquímics significatius que es producen18. Tot i així, s’estan realitzant nous estudis en asma i altres atòpies per tal de millorar el curs de la malaltia.
Diversos autors defensen un altre tipus d’àcids grassos diferents als w-3 per pensar que pogués arribar a un estat d’immunosupressió que deixés a l’individu desprotegit i comprometés el seu estat de salud19. No obstant això, els nostres resultats suggereixen que els àcids grassos w-3 són capaços de potenciar algunes defenses de l’organisme, com ara la immunoglobulina A, l’increment pot estar també potenciat per l’augment d’IL-10 que indueix el canvi d’isotip de les immunoglobulines. En aquest estudi també s’observa l’increment en l’expressió de iNOS en macròfags que suposaria una ajuda davant d’un atac extern.
En la nostra opinió, el resultat més rellevant i nou d’aquest estudi és l’increment de la citocina reguladora IL-10, la qual podria ser considerada responsable de l’efecte antiinflamatori dels àcids grassos w-3, així com de l’antial·lèrgic, ja que el poder immunosupressor d’aquesta citocina podria disminuir les respostes tant Th1 com Th2.
Diversos estudis, com els realitzats en models de pancreatitis experimental20 on es va aplicar nutrició parenteral amb oli de peix, coincideixen que els àcids w-3 indueixen un increment dels nivells d’IL-10. No obstant això, altres autors no observen diferencias21 o fins i tot perceben una disminució de la IL-1022 en altres models. Seria recomanable aprofundir en aquest fet per intentar concretar el paper de la IL-10 en el mecanisme d’acció dels àcids grassos w-3 pel que estudis amb Knock-out per a IL-10 estan previstos realitzar-se en els nostres laboratoris.
el model de dermatitis de contacte emprat en aquest estudi, permet l’observació a simple vista de l’efecte antiinflamatori de la dieta enriquida en àcids grassos w-3. Els valors de l’pes de les orelles i de el contingut d’aigua de les mateixes ja reflecteix el clar efecte amiinflamatorio de l’oli de peix. Que el pes i el contingut d’aigua de les orelles inflamades de ratolins alimentats amb w-3 fos menor era indicatiu que en aquest grup la vasodilatació havia estat menys dràstica i per tant l’edema més lleu. Estudis japonesos confirmen aquests efectes en un mode-ho similar, atribuint en aquest cas l’efecte a l’major poder de l’DHA7. Un estudi semblant, usant èsters de phorbol (TPA) com a agent irritant, també assenyala el poder antiinflamatori, en aquest cas per igual, de DHA i EPA23.
La mesura de MPO posa de manifest una menor infiltració leucocitària amb el tractament d’àcids grassos w-3. La migració de leucòcits des de la microvasculatura fins al focus de la lesió on s’acumulen, és un pas clau característic de la inflamació aguda23,7. Els àcids grassos w-3 semblen posseir un important paper a aquest nivell ja que en aquest grup tot just es percep el fenomen.
Els radicals lliures associats a la inflamació porten a estrès oxidatiu, involucrat en l’agreujament de nombroses malalties. El glutatió és un tripèptid d’elevada capacitat antioxidant a què recorre l’organisme per a la neutralització dels radicals lliures. A les orelles de ratolins on es va induir la dermatitis de contacte dels nivells de glutatió disminueixen significativament a l’afrontar l’estrès oxidatiu generat. No obstant això, en el grup w-3 aquesta disminució està reduïda o compensada ja que a l’ésser el dany menor, el seu efecte antioxidant no és tan requerit. Aquests resultats podrien suggerir que els àcids grassos w-3 augmentin els nivells de glutatió, però, a l’analitzar el contingut de glutatió en el fetge de tots els rosegadors no es van observar diferències. No obstant això, sí que hi ha estudis que relacionen el consum d’àcids grassos w-3 amb un increment en les defenses antioxidantes24,25.
Podem concloure atribuint als àcids grassos w-3 un potent paper antiinflamatori on el augment de la IL-10 sembla ser el factor desencadenant d’una sèrie de modificacions en la resposta immunològica que porten a aquest efecte. La disminució de la proliferació de limfòcits i de les resposta de tipus Th1 i Th2 són algunes d’aquestes modificacions. L’estudi de l’efecte dels àcids w-3 a nivell de vies de transducció de senyals es fa imprescindible per acabar de donar explicació a la gran activitat d’aquests compostos, no només a nivell d’inflamació sinó també a nivell d’al·lèrgia.
Referències
1. Kelley D: Modulation of human immune and inflammatory responses by dietary fatty acids. Nutrition 2001, 17 (7/8): 669-673.
2. Calder PC: n-3 poliinsaturats fatty acids and Inflammation: from molecular biology to the clínic. Lipids 2003, 38: 323-352.
3.Harbige LS: àcids grassos, la resposta immune i autoimmunitat: una qüestió de la essencialitat N-6 i l’equilibri entre N-6 i N-3. Lípids 2003, 38: 323-341.
4. Simopoulos AP: àcids grassos Omega-3 en inflamacions i malalties autoinnummes. J Am Coll Nutr 2002,21 núm. 6: 495-505.
5. PC Calder: Possibilitat de la immunomodulació. Tendències Immunol avui 1996, 19, no 6: 244-247.
6. Yaqoob P: lípids i la resposta immune: del mecanisme molecular a les aplicacions clíniques. CURS OP MOUR NUT METAB CARE 2003, 6: 133-150.
7. Tomobe Yi, Morizawa K, Tsuchida M, Hibino H, Nakano Y, Tanaka Y: Dietètic Docosahexaenoic àcid suprimeix la inflamació i les immunes. LIPIDS 2000, 35, NO. 1: 61-69.
8. Anderson Me: Determinació de glutatió i glutatió disulfur en mostres biològiques. Meth Enzymol 1985, 113: 548-555.
9. Krawisz Ye, Sharon P, Stenson WF: Assaig quantitatiu per a la inflamació intestinal aguda basada en l’activitat de la mieloeroxidasa. Valoració de la inflamació en models de rata i hàmster. Gastroenterologia 1984, 87: 1344-1350.
10. Kelley DS, Warren JM, Simon Va, Bartolini G, Mackey Be, Erickson KL: efectes similars de C9, T11-CLA i T10, CL2CLA sobre funcions de cèl·lules immunitàries en ratolins. Lípids 2002, 37: 725-728.
11. Xaus J, Cardó M, Vellador AF, Soler C. Lloveras J, Celada A: Interferon Gamma indueix l’expressió de P21WAF1 i detencions Cicle de cèl·lules macròfags que impedeix la inducció de l’apoptosi. Immunitat 1990, 2: 103-113.
12. Peterson LD, Thies F, Sanderson P, Newsholme EA, Calder PC: nivells baixos d’àcids eicosapentaenoics i docosahexaenoics imiten l’efecte de l’oli de peix sobre limfòcits de rata. LIFE SCI 1998, 62 (24): 2209-2017.
13. Peterson LD, Jeffery NM, Thies F, Sanderson P, Newsholme EA Y VALDER PC: àcids eicosapentaenoics i docosaahexaenoics alterar la melsa de metalls de la rata de la composició de les àcids grassos i la producció de prostaglandin E2, però tenen efectes diferents sobre la funció de limfòcits i la cèl·lula. Lípids 1998, 33 (2): 171-180.
14. Thies F, Neve-von-Caron G, Powell JR, Yaqoob, P, Newsholme EA, Calder PC: suplements dietètics amb àcid gamma-linolènic o l’oli de peix disminueix la proliferació de limfòcits en humà més gran. J Nutr 2001, 131 (7): 1918-1927.
15. Kew S, Benerjee T, Minihane Am, Finnegan Ye, Muggli R, Albers R, Williams CM, Calder PC. La manca d’efecte dels aliments enriquits amb àcids grassos n-3 derivats de la planta ormarina sobre les funcions immunes humanes. Am J Clin Nutr 2003, 77 (5): 1287-1295.
16. Trebble TM y Cols.: PRODUCCIÓ PRODUCCIÓ DE PROSTAGLANDIN E2 i Funció de cèl·lules T Després de la suplementació de peixos: resposta a la cosupplementació antioxidant. Am J Clin Nutr 2003, 78 (3): 376-382.
17. Endres S y cols.: L’efecte de la suplementació dietètica amb àcids grassos N-3 sobre la síntesi de la interleucina-1 i el factor de necrosi tumoral per cèl·lules mononuclears. N Engl J Med 1989, 320 (5): 265-271.
18. Calder PC, Yaqoob P, Thies F, Wallace Fa Y milles EA: àcids grassos i funcions de limfòcits. Brit J Nutr 2002, 87 Supl. 1, S31-S48.
19. Puertollano Ma, de Pablo M, Álvarez de Cienfuegos G: Propietats antioxidants de N-Acetcine-Cistcó No milloren la resistència immune dels ratolins alimentats diàries alimentats a Listeria Monocytogenes Infection. Clin Nutr, 2003, 22 (3): 313-319.
20. Foitzik T, Eibl G, Schneider P, Wenger Fa, Jacobi CA, Buhr HJ: Omega-3 Splement d’àcids grassos augmenta les citocines antiinflamatòries i atenua les seqüeles de malalties sistèmiques en pancreatis experimentals. Parenter Enteral Nutr, 2002, 26 (6): 351-356.
21. Wallace Fa, Miles EA, Calder PC: comparació dels efectes de l’oli de llinosa i les diferents dosis d’oli de peix a la funció de cèl·lules mononuclears en subjéss humans sans. BR J Nutr 2003, 89 (5): 679-681.
22. Hayashi y co .: Efectes de la intravenosa Omega-3 i l’emulsió de greixos Omega-6 sobre la producció de citocina i la sensibilitat de la hipè de tipus retardada en rates cremades que reben la nutrició total parental. J Parenter Enteral Nutr 1998, 22 (6): 363-367.
23. Raerlerstorff D, Pantze M, Bachmann H, Moser U: Propietats antiinflamatòries dels àcids docosahexaenoics i eicosupentaenoics en la inflamació de l’orella del ratolí induïda pel phorbol-ester. Int Arch Allergy Immunol 1996, 111 (3): 284-290.
24. Jolly Ca, Muthukumar A, Avula CP, Troyer D, Fernandes G: La vida útil és poc gravada en els ratolins autoimmunes (Nzbxnzw Nzbxnzw) f (L) alimentat una dieta enriquida amb (N-3) àcids grassos. J Nutr 2001, 131 (10): 2753-60.
25. Demoz A, Willumsen N Y Berge RK: àcid eicosapentaenoic a la dosi hipotrigliceridèmica millora la defensa antioxidant hepàtica en ratolins. Lípids 1992, 27 (12): 968-971.