ORIGINAL
Alguns aspectes de la dinàmica poblacional de l’sorell Caranx hippos (pisces: Carangidae) en Boques de Cendra, Carib colombià
Some aspects of the population dynamics of the crevalle jack Caranx hippos (pisces: Carangidae) in Boques de Cendra, colombian Caribbean
a
1Ministerio d’Agricultura i Desenvolupament Rural, Bogotà, Colòmbia.
2Universidad de l’Magdalena, Programa d’Enginyeria Pesquera, Grup de Biodiversitat i Ecologia Aplicada, Santa Marta, Magdalena, Colòmbia.
3Corporación Colòmbia Internacional (CCI), Barranquilla, Atlántico, Colòmbia.
* Correspondència: [email protected]
Rebut: setembre de 2009; Acceptat: Juliol de 2010.
Resum
Objectiu. Aquest estudi va tenir com a propòsit avaluar aspectes biològics i la dinàmica poblacional de l’Sorell (Caranx hippos) al sector de Boques de Cendra, Carib colombià. Materials i mètodes. Entre juny de 2005 i juny de 2006, es van recol·lectar dades de freqüències de longituds comercials per a determinar la relació longitud forquilla-pes (LH-W), la talla mitjana de maduresa sexual (L50%), els paràmetres de creixement (K, L ∞ i t0), les taxes de mortalitat (M, F i Z) i d’explotació (e). Es va comptar amb 1151 individus, dels quals 264 es van usar per a l’anàlisi biològic. Resultats. La relació LH-W va diferir entre sexes. Es va estimar una L50% en 63.6 cm de LH. Els paràmetres de creixement van presentar els següents valors: L∞ = 91 cm de LH, K = 0.38 any-1 i to = 0.32 any. El valor de l’índex d’acompliment va tenir un rang entre 3.33 i 3.73. La taxa de mortalitat total (Z) va ser de 1.85 any-1, la mortalitat per pesca (F) de 0.72 any-1 i la natural (M) en 1.13 any-1. L’estat d’explotació (E) va ser de 0.30 any-1. Conclusions. Segons la taxa d’explotació, el Sorell no presenta sobreexplotació en Boques de Cendra. No obstant això, a l’comparar la talla mitjana de captura amb la L50%, l’espècie està sent capturada amb els principals arts de pesca abans que el 50% de la població aconsegueixi madurar. Es proposen pautes per al disseny d’un sistema de maneig pesquer que garanteixi la conservació i l’aprofitament sostenible d’aquest recurs.
Paraules clau: Pesca, reproducció, Caranx hippos, Carib, Colòmbia. (Font: AIMS).
Abstract
Objetive. To evaluate the biological aspects and the population dynamics of the Crevalle jack (Caranx hippos) from Boques de Cendra, colombian Caribbean. Materials and methods. Between June 2005 and June 2006 length frequency data were collected to determini fork length-weight relationship, pixen maturity size (L50%), Von Bertalanffy ‘s equation growth parameters (K, L∞ i t0), mortality (M, F i Z) and exploitation (I) rates. A total of 1151 individuals were Sampled, and 264 were used for Biologic analysis. Results. Fork length-weight relationship showed differences between genders. L50% was estimated in 63.6 cm FL. Growth parameters values were: L∞ = 91 cm FL, K = 0.38 year-1 i to = 0.32 year. The comparative growth index (Φ ‘) values ranged from 3.33 to 3.73. Total mortality rate (Z) was estimated as 1.85 year-1, fishing mortality (F) es 0.72 year-1 and natural mortality (M) as 1.13 year-1. Exploitation rate (E) was determined as 0.31 year-1. Conclusions. The exploitation rate indicated that Crevalle jack from Boques de Cendra was not overexploited; before 50% of the population reach its sexual maturity. We propose a framework for a Fishery management system, assuring the conservation and suitable use of this resource.
Key words: Fishing, reproduction, Caranx hippos, Carib, Colòmbia. (Source: AIMS).
Introducció
La família Carangidae és un dels grups de peixos de major importància econòmica en les pesqueres tropicals i subtropicals de l’món (1, 2). Dels desembarcaments provinents de la pesca artesanal al departament de l’Atlàntic, el Sorell Caranx hippos (Linneo 1766) és una de les espècies de major demanda (3). Representa el 47% en les captures desembarcades anualment i el 70% dels ingressos econòmics de la població de pescadors, sent capturada principalment amb tremalls i en alguns casos amb hams i boliches (3). Tot i això, hi ha poca informació detallada sobre els aspectes biològics i poblacionals que permetin dilucidar l’estat d’explotació pesquera en el qual es troba la seva població a la desembocadura del riu Magdalena.
Les estimacions dels paràmetres de creixement, les taxes de mortalitat i d’explotació són part important en l’estudi de la biologia dels peixos d’interès comercial, ja que permeten determinar els nivells d’explotació i al seu torn proporcionen eines per a la implementació de polítiques d’administració i control per a un adequat aprofitament (4-6). L’objectiu d’aquest treball va ser determinar la relació LH-W, la talla mitjana de maduresa sexual, els paràmetres de creixement, les taxes de mortalitat i d’explotació de C. hippos en Boques de Cendra per conèixer l’estat d’explotació en aquest sector i fonamentar les bases científiques per proposar pautes orientades a el disseny de sistemes de maneig pesquer que condueixin a una explotació sostenible d’aquest important recurs.
Materials i mètodes
Àrea d’estudi. Va comprendre l’estuari del riu Magdalena en el Mar Carib, sector denominat Boques de Cendra (11 ° 06 ‘- 11 ° 08’ Nord; 74 ° 50 ‘- 74 ° 52’ Oest; Figura 1). Aquest sector es troba localitzat a nord de l’departament de l’Atlàntic (3). El clima es caracteritza per la presència de dos períodes climàtics que afecten l’activitat dels pescadors (7); el sec que comença a principis de desembre i acaba a finals d’abril sense precipitacions i una disminució de l’cabal del riu Magdalena amb una mitjana de 4068 m3 s-1 (7, 8). El període plujós inicia al maig i finalitza a principis de desembre amb disminució dels vents i alta nuvolositat (7, 9) i és quan es presenten les majors descàrregues d’aigua amb una mitjana de 10287 m3 s-1 (8).
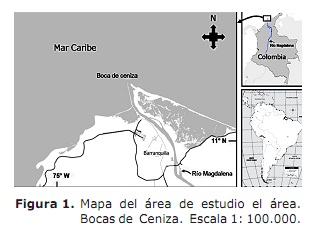
Boques de Cendra és un estuari de falca salina, on el seu funcionament respon a la interacció entre l’ona de marea i el aportació fluvial dins dels contorns imposats per la geometria de la mateixa estuari (9). El sector està influït per dos corrents: la provinent de l’Carib amb sentit oest i el corrent de Panamà amb direcció est (9). En aquesta àrea, l’activitat pesquera és de tipus artesanal i compta amb prop de 250 pescadors que operen els arts de pesca tremalls, hams, palangres i bolig. Les captures estan representades per espècies marines, així com d’espècies estuarinas. Es destaquen Macrodon ancyclodon, sorella del sud, C. hippos, Scomberomorus brasiliensis, Ariopsis sp., Bagre spp. i Arius props (3).
Fase de camp i laboratori. Entre juny de 2005 i juny de 2006 es van realitzar mostrejos durant cinc dies de cada mes per registrar freqüències de longituds de les captures comercials desembarcades al barri Les Flors, Barranquilla. Les captures van ser realitzades amb hams de calibre 7-10 i amb boliches i tremalls entre 8.89 i 20.32 cm de mida de malla. Cada individu va ser sexat i pesat (sense eviscerar) amb una balança de precisió de 0.1 g. Atès que no tots els peixos van arribar amb les aletes cabals completes, es va usar la longitud forquilla. Els mesuraments es van realitzar a el límit inferior, utilitzant un ictiómetro amb 0.1 cm de precisió (3). Per a l’anàlisi biològic es van realitzar observacions macroscòpiques de les gònades per determinar el sexe i estat de maduració gonadal, prenent com a referència la classificació qualitativa de Vazzoler (10), la qual consisteix en quatre estats: indiferenciats; immadurs (I); en maduració (II); madurs (III) i desovados (IV).
Relació longitud-pes (LH-W). Es va determinar la relació longitud-pes per sexe a través de l’equació potencial W = aLHb, on W és el pes total, a i b són constants de la regressió potencial i LH és la longitud forquilla. Per determinar diferències en la relació LH-W entre sexes, es va realitzar una anàlisi de covariància (ANCOVA), on la variable dependent va ser W, la variable independent va ser LH i el factor el sexe. Previ a l’anàlisi es van avaluar els supòsits de normalitat dels residus (prova de Kolmogorov-Smirnov), homogeneïtat de variàncies (prova de Bartlett), relació lineal entre la variable W i LH (anàlisi de correlació), i homogeneïtat de les pendents de la relació entre W i LH.
Talla mitjana de maduresa sexual (L50%), composició de la captura per talles i talla mitjana de captura (TMC). Per avaluar l’impacte de la pesqueria sobre la biologia de C. hippos, es va calcular L50%. Es van considerar aquells exemplars les gònades es van observar en els estats III i IV. El càlcul es va basar en el model logístic que relaciona la proporció d’individus madurs amb la longitud (L; 5):
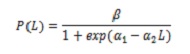
on, β = 1; α1 i α2 són els paràmetres de la regressió.
Per a estimar L50% es va usar la relació -α1 / α2. Per ajustar el model es va utilitzar el mètode de regressió no lineal per mínims quadrats, usant l’algoritme iteratiu de Newton.
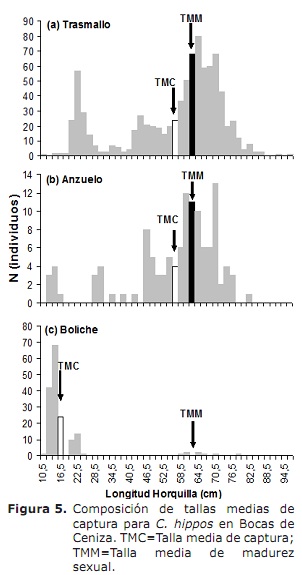
La composició de la captura per talles va ser usada per calcular la TMC i els paràmetres de creixement.Les dades van ser agrupats mensualment en marques de classe cada dos centímetres. La relació entre la freqüència relativa i el nombre total d’individus es va emprar per calcular la TMC usant el procediment proposat per Sparre i Venema (5); posteriorment, es va calcular una TMC per a cada art de pesca observat. Les TMC van ser comparades amb la L50% per avaluar l’impacte de la pesca sobre la biologia de l’espècie.
Paràmetres de creixement. Les dades de longitud van ser agrupats mensualment en marques de classe d’1 cm. Els paràmetres de creixement es van estimar mitjançant tres rutines contingudes en el paquet computacional FISAT versió 1.1 (4): l’anàlisi de freqüència electrònica (ELEFAN I), el de composició de longituds d’Shepherd (SLCA; 11) i el mètode de projecció de matrius (PROJMAT; 12). La rutina ELEFAN I treballa amb l’equació de creixement estacionalitzada de von Bertalanffy (ELEFAN I), sent la talla expressada en funció de l’edat (5):
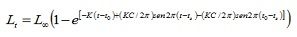
on, Lt és la longitud a l’edat t; L∞ la longitud asimptòtica; K és la constant de creixement, to és el paràmetre de condició inicial, el qual determina el punt hipotètic en el temps en què el peix té una talla igual a zero; C és el paràmetre d’amplitud de l’oscil·lació de el creixement; ts és la part de l’any en què la taxa de creixement és mínima.
La rutina SLCA estima el creixement independent de l’nombre i posició de les modes en la mostra, basat en l’ajust de la funció cosinus:
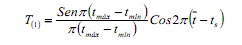
on, tmáx i Tmín són edats corresponents als límits superior i inferior d’un determinat interval de classe; és l’edat mitjana i ts és la fracció de l’any en què va ser presa la mostra (6).
L’PROJMAT projecta una mostra de distribució de freqüències de longituds de temps ta temps t + 1, fent servir els valors de K i L∞. Les freqüències projectades van ser comparades amb les observades per la suma de les diferències de quadrats, repetint el procés fins a obtenir els millors estimatius (6).
Tenint en compte que no és possible obtenir una estimació de to a partir de les freqüències de longituds (13,14), aquest paràmetre va ser estimat a partir de l’equació empírica de Pauly (15):
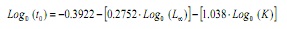
on, to és l’edat hipotètica a la qual comença a créixer l’animal, L∞ la longitud asimptòtica i K és la constant de creixement.
Per a comparar les estimacions de creixement entre mètodes, es va utilitzar l’índex d’acompliment de creixement Φ ‘(16). Addicionalment, aquests valors van ser comparats amb les estimacions d’altres poblacions de C. hippos documentats en la literatura. Aquest índex es va estimar a partir de l’expressió:
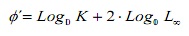
on, K i L∞ són els paràmetres de creixement.
Atès que el valor estimat de L∞ en aquest estudi es va basar en LH i els documentats en la literatura en longitud total (LT), per a propòsits de fer comparacions entre els valors, es va fer una regressió lineal simple entre LT (dependent) i LH (independent) per fer la conversió de LH a LT. Per això es va obtenir el següent model lineal amb dades d’aquest estudi:
LT = – 0,4387 + (1.2335 * LH)
A partir de les freqüències de LH de les captures comercials, es va procedir a estimar les taxes de mortalitat total (Z), natural (M) i per pesca (F) de la població de C. hippos. Z es va estimar a partir de la corba de captura linealizada convertida a talles (17):
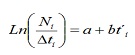
on, Ni és el nombre d’individus capturats pertanyents a la classe de longitud i; t’i és l’edat relativa de la classe i; Δti és el temps mitjà requerit per individu per al seu creixement a través de la classe i.
M va ser estimada utilitzant l’equació empírica de Pauly (17):
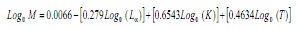
on, L∞ i K són els paràmetres de l’equació de von Bertalanffy i T és la temperatura mitjana anual de l’aigua costanera superficial en l’àrea d’estudi, la qual es va considerar en 28.2 ° C.
la taxa de mortalitat per pesca i la taxa d’explotació van ser estimades a partir de:
Per a realitzar les estimacions dels paràmetres es va assumir un creixement estacionalizado de C. hippos, atès que a través d’l’any succeeixen episodis ambientals en l’àrea d’estudi que podrien estar influint en el creixement dels peixos. Tal és el cas de les condicions estacionals de l’cabal del riu Magdalena, les quals poden influir en les condicions físiques, químiques i biològiques en la seva desembocadura (8). A això se li suma, la influència dels vents Alisios en el sector de Boques de Cendra, ja que quan estan en marxa i segons la seva intensitat, també canvien les condicions oceanogràfiques de el sector (18). De la mateixa manera, els coneixements biològics de l’espècie indiquen que C.hippos és una espècie migratòria (1), a partir de la qual cosa també es pot inferir estacionalitat en el creixement. Amb base a l’anterior es van seleccionar els valors trobats per ELEFAN I per estimar les taxes de mortalitat de C. hippos en aquest estudi.
Amb el propòsit de quantificar l’efecte de les mostres mensuals en els paràmetres de creixement i obtenir els valors de precisió dels mateixos, es va aplicar la tècnica de remostreig Jackknife (6). Aquesta consisteix a estimar les n mostres mensuals de freqüència de talles i seqüencialment són omeses per obtenir n combinacions de cadascun dels paràmetres. Per precisar aquesta estimació es va usar el Error Percentual (EP) i el Coeficient de Variació (CV), utilitzant les següents fórmules: a
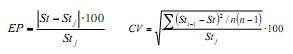
on, St és el paràmetre de creixement estimat utilitzant la base de dades anual completa i STJ és l’estimador mitjana derivat de les estimacions successives de Jackknife; Sti-1 és el paràmetre estimat a l’ometre el mes i, i n és la mida de la mostra anual (n = 12).
Resultats
Durant el període d’estudi es van recol·lectar 1151 individus, dels quals 868 van ser capturats amb tresmall, 120 amb ham i 167 amb bolig; presentant un rang de talla comprès entre 10.5 i 96.5 cm de longitud forquilla. Per a l’anàlisi dels aspectes biològics estudiats es va comptar amb 264 individus, sent 180 mascles i 84 femelles, amb un rang de talla entre 25 i 96.5 cm de LH i un pes total entre 281.5 i 10500 g.
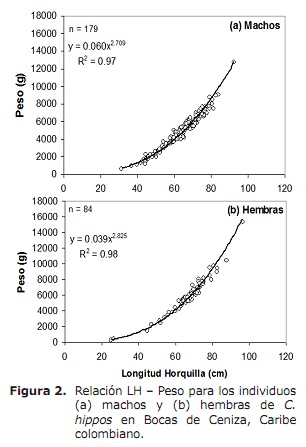
Relació longitud-pes (LH-W). L’anàlisi va revelar que entre sexe la relació LH-W va diferir significativament, observant-se que les femelles pesen més que els mascles a una mateixa longitud (ANCOVA: F1, 260 = 21.5; p < 0.05 ). Per tant, la relació LH-W a mascles va ser W = 0.060LH2.709 (p < 0.05) i per femelles W = 0.039LH2.825 (p < 0.05) (Figura 2). Els valors de b resultants de les regressions per a tots dos sexes van ser significativament menors de 3, indicant un creixement minorant (prova t; p > 0.05; 15).
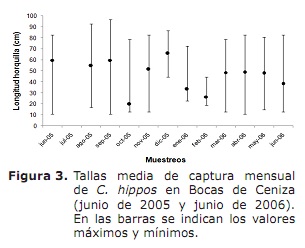
Composició per talles, talla mitjana de maduresa sexual (L50%) i talla mitjana de captura (TMC). De manera general, les talles mínimes de captura es van registrar a l’octubre, novembre de 2005 i març, abril i juny de 2006; mentre que les màximes van ser freqüents durant tot el període d’estudi, excepte per a l’octubre, gener i febrer (Figura 3).
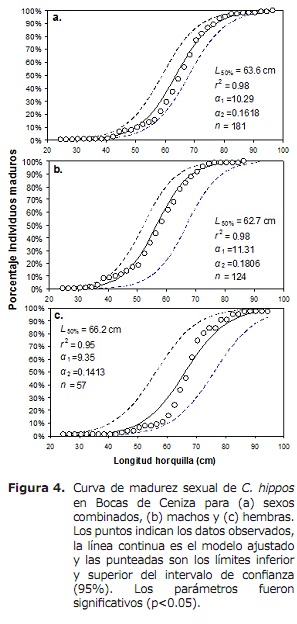
La L50% per sexes combinats va ser 63.6 cm de LH (IC a el 95%: 59.0-68.4 cm). Per al cas de mascles i femelles van ser 62.6 cm (IC a l’95%: 58.3-70 cm) i 66.2 cm (IC a l’95%: 56.1-76.2 cm), respectivament (Figura 4). La TMC global va ser estimada en 50.3 cm de LH. Per al cas de tremalls, hams i boliches es van estimar en 56.0, 56.2 i 17.8 cm, respectivament (Figura 5a, 5b i 5c), per sota de la L50%.
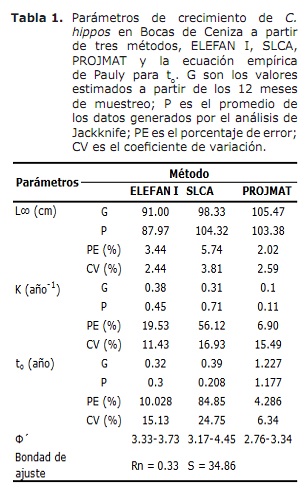
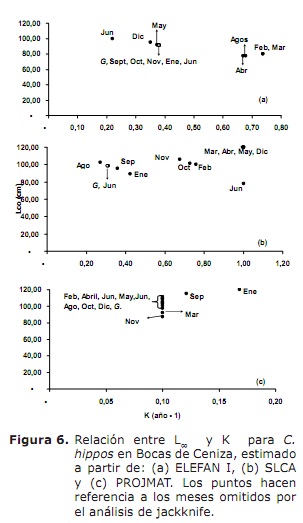
Creixement. Els resultats de L∞ i K proporcionats pels mètodes ELEFAN I, SLCA i PROJMAT es presenten a la taula 1. S’observa que aquests tres mètodes van subministrar diferents valors per a cada un dels paràmetres de creixement. L’anàlisi de Jackknife va mostrar que hi ha efecte de les dades sobre els diferents paràmetres de creixement, especialment per SLCA i PROJMAT. Per al cas de ELEFAN I, L∞ va variar de 77.4 a 100 cm, mentre que els valors de K van estar entre 0.20 i 0.75 any-1 (Figura 6). Amb SLCA es van obtenir els valors més alts de L∞ i K amb valors entre 78.25 i 120 cm i entre 0.3 i 1.0 any-1 respectivament. D’acord amb els valors llançats per PROJMAT, es va observar menor dispersió per a K fluctuant entre 0.10 i 0.17 any-1. El contrari va passar per L∞, on els valors van variar entre 87 i 120 cm, sent aquest últim major a l’observat en els estimadors de creixement anteriors. Els valors de K estimats mostrar diferències entre mètodes, sent els més alts els calculats per ELEFAN I i SLCA (Taula 1). El valor obtingut per l’edat hipotètica a la qual comença a créixer l’animal (to) mitjançant l’equació empírica de Pauly (20) va ser de 0.32 any, amb un coeficient de variació de 13.92% i un percentatge d’error (PE) de 10.02 . L’anàlisi de l’índex d’acompliment ǿ mostrar valors similars entre ELEFAN I (3.33-3.73) i SLCA (3.17-4.45), el que no va passar per PROJMAT que va mostrar valors inferiors als anteriors (2.76-3.34). Tot i així, la menor variació es va presentar per ELEFAN I, seguit per PROJMAT i SLCA, sent aquest últim el que major rang presentar. Tenint en compte el coeficient de variació (CV) de cada un dels paràmetres, es va trobar que els estimatius amb ELEFAN I de l’anàlisi de Jackknife tendir a proporcionar menors valors per L∞, a diferència de SLCA i PROJMAT els quals van aconseguir valors més alts (Taula 1).El resultat de l’PE obtingut per K amb PROJMAT va ser menor a l’trobat per ELEFAN I i SLCA. D’acord amb això, el mètode que va llançar els estimatius més creïbles dels paràmetres de creixement va ser PROJMAT, seguit de ELEFAN I i finalment SLCA.
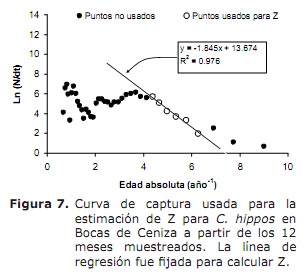
Mortalitat i estat d’explotació . Amb base en els següents paràmetres de creixement: K = 0.38 any-1; L∞ = 91 cm LH; to = 0.32 any; oscil·lació estacional en el patró de creixement (C) de 0,5 i una taxa de creixement mínima (WP) de 0.3, es va estimar una taxa de mortalitat total (Z) de 1.76 any-1 (Figura 7), oscil·lant entre 1.38 i 2.22 any -1 (a partir de l’anàlisi de Jackknife).
La mortalitat natural (M) va ser de 1.12 any-1, variant entre 1.10 i 1.14 any-1. La taxa de mortalitat per pesca (F) es va estimar en 0.63 any-1, presentant un rang de variació entre 0.2 i 1.06 any-1. Els valors de la relació F i Z per a estimar la taxa d’explotació (E), va presentar un valor màxim de 0.5 any-1, presentant-una mitjana de 0,30 any-1.
Discussió
Relació longitud-pes (LH-W). El concepte de creixement implica canvis relatius a la talla i el pes dels individus (19), i la seva relació té diversos usos en la investigació pesquera. Per exemple, és útil en els models d’anàlisi d’estoc pesquers per estimar la biomassa d’una població; per conèixer la condició d’un peix; per comparar la història de vida de certes espècies entre regions; i per determinar el tipus de creixement que solen presentar els peixos (20). C. hippos, presenta un creixement en pes de tipus al·lomètric minorant (< 3; 15), indicant que presenta taxes de creixement diferencial per a diferents parts de el cos, a on la longitud augmenta a una taxa més gran que el pes.
Els resultats indiquen que aquesta condició va ser més evident per a mascles que per femelles, atès que aquestes últimes, tot i presentar un creixement al·lomètric minorant, s’aproximen a tenir un creixement isomètric. Això va ser concordant amb el resultat de la ANCOVA, la qual va indicar que a una mateixa LH ha diferències en el pes entre sexes. Les femelles d’aquesta espècie presenten major pes i alçada corporal que els mascles. D’altra banda, és important ressaltar que pel fet que les mostres van ser recollides durant un any, la relació LH-W en aquest estudi no representa una particular època de l’any. En aquest sentit, els valors de b podrien considerar com un valor mitjà anual per a l’espècie (20). Els resultats d’aquesta investigació van ser similars als documentats per a l’espècie en diferents àrees geogràfiques. Per exemple, es van documentar valors de b per sexes combinats de 2.73 al Sud de la Florida (21), de 2.85 a l’Índia (1) i de 2.91 en el Golf de Salamanca, Colòmbia (22).
Creixement, mortalitat i estat d’explotació. Amb base en els paràmetres de creixement obtinguts per ELEFAN I, la longitud màxima estimada per a C. hippos en Boques de Cendra va ser de 91 cm LH que correspon a 111.81 cm de LT. Si aquest últim és convertit a edat correspon a 14 anys, aconseguida amb una taxa de creixement K de 0.38 any-1, suggerint que C. hippos és una espècie longeva (17). D’altra banda, tenint en compte la corba de creixement, aquesta espècie va créixer ràpidament fins a l’edat de 5 anys. Aquesta dada coincideix per a altres caràngids, com el de Trachinotus falcatus, el qual aconsegueix una talla màxima de 90 cm i presenta també un creixement màxim de 5 anys, a partir de la qual s’inicia la fase estacionària del seu creixement (2).
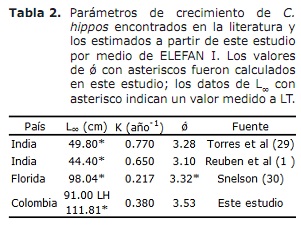
d’altra banda, els valors de L∞ estimat per a C. hippos en aquest estudi van ser superiors als documentats en la literatura; mentre que els de l’índex d’acompliment ǿ revisats en la literatura van mostrar valors molt propers a l’estimat en aquest estudi (Taula 2). Prenent aquest índex com un criteri de comparació entre els valors dels paràmetres de creixement obtinguts i tenint en compte que C. hippos és una espècie migratòria (23), es pot suposar que el creixement d’aquesta espècie és concordant en el descrit en diferents àrees de la seva distribució mundial (Taula 2).
El patró anual de creixement de C. hippos va mostrar oscil·lacions (C = 0.5 ) amb una taxa mínima de creixement en el mes de setembre (WP = 0.3). Aquest patró pot estar influït per un dels aspectes més característics en el creixement dels peixos: la variabilitat observada entre individus d’una població i entre les poblacions d’una espècie, la qual està directament relacionada amb el medi ambient (24). Per exemple, la variabilitat en les condicions fisicoquímiques de l’aigua (24) i la qualitat i quantitat d’aliment (19) condicionen la migració d’una espècie, la qual abasta desplaçaments estacionals entre les zones de cria, alimentació i fresa (15, 25).
Amb base en l’anterior i tenint en compte les freqüències de longituds observades durant el període d’estudi, les quals també indiquen la possible presència d’estacionalitat en el creixement (13), es pot inferir que el creixement de C. hippos és estacional. Pel que fa a la taxa mínima de creixement, aquest esdeveniment pot estar donat possiblement per l’etapa reproductiva de l’espècie, ja que per a setembre es va observar un increment en el nombre d’individus madurs (en estadis III).
D’altra banda, la relació entre la taxa de mortalitat per pesca (F) i mortalitat total (Z) de 0.3, indica que la població en l’àrea de Boques de Cendra no està corrent risc de ser sobreexplotada , sota la premissa que una població aconsegueix el seu rendiment òptim quan la fracció de morts causada per F està per sobre de 0.5 (6,13). No obstant això, és important ressaltar que quan L50% va ser comparada amb la TMC, C. hippos presentar una major pressió sobre la població d’adults, freqüentment capturada amb tresmall i ham. La situació empitjora amb l’ús de el bolig, el qual no només afecta els adults, sinó també als joves, el que no permet la renovació natural d’aquest recurs pesquer.
Proposta per al disseny d’un sistema de maneig pesquer. Partint de la base que la sobrepesca pot tenir lloc en tres formes diferents: (i) en relació a l’creixement; (Ii) pel que fa a l’reclutament; i (iii) sobrepesca que afecta l’ecosistema (15). En aquest sentit, la pesca d’C. hippos en el sector de Boques de Cendra es pot classificar dins el primer ítem, la qual té lloc quan els individus són capturats abans que arribin a la talla necessària per garantir la renovació biològica de la població. L’anterior s’ha convertit en un problema comú per a la majoria de les espècies comercials, i ve sent documentat per a moltes espècies costaneres al Carib colombià.
Tot això, fa necessari que s’inverteixin esforços per dissenyar estratègies de maneig per a C. hippos a la pesquera de Boques de Cendra, de tal manera que siguin consistents amb l’enfocament precautori per a la pesca responsable d’aquest recurs (27). Caddy (27) suggereix que les estratègies han de ser aplicades dins d’un marc de redundància de maneig on tal marc podria consistir en una barreja de mesures que involucri criteris de selectivitat d’arts, vedes temporals i l’augment de les talles mínimes de captura. En aquest sentit, tenint en compte la L50% com un punt de referència límit (28), aquí es recomana que la talla mínima de captura d’aquest recurs hagi de fixar-se en 64 cm de longitud forquilla. Així mateix, en tots els arts de pesca utilitzats es proposa incrementar les mides de malla per sobre dels 8.89 cm i usar calibres d’hams dirigits a capturar peixos més grans. En les condicions actuals aquests arts de pesca estan afectant directament a la fracció d’individus joves que ingressen a la primera fase de maduresa sexual. Amb les mesures anteriors, complementades amb altres variables com la captura, l’esforç de pesca i la captura per unitat d’esforç, així com també factors de socials i econòmics, s’espera que C. hippos es capturi en una major proporció d’individus per sobre de la L50% per garantir la sostenibilitat d’aquest recurs a través del temps. En aquest sentit, és important tenir en compte la condició multiespecífica i multiflota de la pesquera i per tant, es recomana un procés de concertació entre la comunitat de pescadors, els comerciants i les entitats pertinents per arribar a acords dirigits a una explotació responsable de l’ recurs.
en conclusió, el Sorell no presenta sobreexplotació en Boques de Cendra. No obstant això, a l’comparar la talla mitjana de captura amb la L50%, l’espècie està sent capturada amb els principals arts de pesca abans que el 50% de la població aconsegueixi madurar. Es proposen pautes per al disseny d’un sistema de maneig pesquer que garanteixi la conservació i l’aprofitament sostenible d’aquest recurs.
Agraïments
A la comunitat de pescadors, als membres de ASOPESCAR i COOPEZ. Als germans Vila i a la peixateria El Nou Tambo per la seva col·laboració en la coordinació d’activitats i subministrament d’informació de camp. Als professors G. Lozano, L.C. Gutiérrez, O. Galvis, J. Blanc i E. Viloria.
a
Referències
1.Reuben, S, HM Kasim, S Sivakami, PN Radhakrihnan, KN Kurup, M Sivadas, A Noble, KVS Nair, Raje SG. Fishery, biology and estoc assessment of Carangid resources from the Indian siguis. Indian J Fish 1992; 39 (3,4): 195-234.
2.Crabtree, R, PB Hood, Snodgrass D. Age, grouth and reproduction of permit (Trachinotus falcatus) in Florida waters. Fish Bull 2002; 100: 26-34.
3.Rivera R, Solano, G.Diagnòstic de l’activitat pesquera artesanal a la zona costanera de l’departament de l’Atlàntic, Carib colombià. Barranquilla, Colòmbia: INCODER; 2005.
4.Gayalino, FC, P Sparre, Pauly D. The FISAT User Guide. FAO – ICLARM Stock Assessment Tools. Roma: FAO; 1993.
5.Sparre, P., Venema S. Introducció a l’avaluació de recursos pesquers tropicals. Roma: FAO; 1995. Document Tècnic de Pesca: 306/1.
6.Rueda, M, Sants-Martínez A. Population dynamics of the striped variada eugerres plumieri from the Pantà Gran de Santa Marta, Colòmbia. Fish Res 1999; 42: 155-166.
7.Blanco, JA, EA Viloria, Narváez BJC. ENSO and interannual Salinity changes in the Pantà Gran de Santa Marta coastal lagoon system, colombian Caribbean. Estuar Coast Shelf Sci 2006; 66: 157-167.
8. Restrepo, JD, Kjerfve B. Magdalena river: interannual variability (1975-1995) and revised water discharge and sediment load estimates. J Hydrol 2000; 235 (1-2): 37-149.
9.Acevedo A, Gutiérrez ML. Alguns aspectes reproductius de l’toyo Rhizotrionodon porosos (1831) (Elasmobranchii: Carcharinidae). En el sector de Boques de Cendra, Carib colombià. . Barranquilla, Colòmbia: Universitat de l’Atlàntic, Facultat de Ciències Bàsiques; 1999.
10.Vazzoler, EAM. Manual de mètodes per estudos biològics de Populaçoes de Peixes: reproducao i crescimento. Brasília: CNPQ; 1982.
11.Sheperd, JG. A weakly parametric method for the analysis of length composition data. In: Pauly, D, Morgan GP editors. Length-based Methods in Fisheries Research. Manila, ICLARM Conference Proceedings 13. 1987.
12.Leslie, PH. The use of matrius in certain population mathematics. Biometrika 1945; 35: 213-245.
13.Gulland, JA, Rosemberg AA. Examen dels mètodes que es basen en la talla per avaluar les poblacions de peixos. Roma: FAO; 1992.
14.Tordecilla-Petro G, S Sánchez-Banda, Olaya-Nieto CH. Creixement i mortalitat de l’Moncholo (HOPLIAS MALABARICUS) en el Pantà Gran de Lorica, Colòmbia. Rev MVZ Còrdova 2005; 10 (2): 623-632.
15.Pauly, D. Alguns mètodes simples per a l’avaluació de recursos pesquers tropicals. Roma: FAO; 1983.
16.Pauly, D, Munro JL. Onze more on the comparison of growth in fish and invertebrates. Fishb 1984; 01:21.
17.Pauly, D. Fish population dynamics in tropical waters; a manual for use with programmable Calculators. ICLARM Stud Rev 1984; 8: 235.
18.Blanco, JA. Les variacions ambientals estacionals en les aigües costaneres i la seva importància per a la pesca a la regió de Santa Marta, Carib Colombià. . Santa Marta-Bogotà. Universitat Nacional de Colòmbia, Facultat de Ciències Bàsiques; 1988.
19.Abaunza L, Karlou-Riga C, Murta A, Eltink W, García Santamaría MT, Zimmermman C, Hammer C, et al. Growth and reproduction of horse mackerel, Trachurus trachurus (Carangidae). Rev Fish Biol Fish 2003; 13: 27-61.
20.Froese, R. Cube law, condition factor and weight-length relationships: history, meta-analysis and recommendations. J Appl Ichthyol 2006; 22: 241-253.
21.Bohnsack, JA, Harper D’. Length-weight relationships of selected marine reef fishes from the Southeastern United States and the Caribbean. NOAA Technical Memoràndum; 1988. NMFS-SEFC-215.
22.Duarte, LO, CB García, I Moreno, G Melo, P Navaixes, N Sandoval, et al. Atles demogràfic dels peixos demersals de golf de Salamanca, Carib colombià: dinàmica poblacional, distribució, alimentació i reproducció. . Versió 1. Santa Marta: INVEMAR-UNAL-COLCIENCIAS; 1999.
23.Honebrink, R. A review of the biology of the family Carangidae whith emphasis on species found in Hawaiian waters. Honolulu: Division of Acuatic resources; 2000.
24.Macer, CT. Some aspects of the biology of the horse mackerel (Trachurus trachurus (L)) in waters around Britain. J Fish Biol 1977; 10: 51-62.
25.Harden, FR. Fish migration. London: Arnold; 1968.
26.Narváez B, JC, F Herrera, Blanco J. Efectes dels arts de pesca sobre la mida dels peixos en una pesquera artesanal de el Carib colombià. Bol Invest Mar Cost 2008; 37 (2): 163-187.
27.Caddy, JF. Fisheries management in the twenty-first century: will new paradigms apply ?. Rev Fish Biol Fish 1999; 9: 1-43.
28.Caddy, JF. Limit reference points, traffic lights, and Holistic approaches to fisheries management with minimal estoc assessment input. Fish Res 2002; 56 (2): 133-137.
29.Torres, FJR, MBTM Norizam, LR Garcés, Silvestre GT. Compilation of population parameters of fishes species commonly caught in trawls in south and southeast Àsia. Manila: World fish center; 2001.
30.Snelson, FF. Biological Studies of crevalle jack (Caranx hippos) in Florida. St. Petesburg: Florida Marina Reseach Institute; 1992.